Ответы на экзамен по анатомии. ОТВЕТЫ НА ЭКЗАМЕН ПО АНАТОМИИ. Экзаменационные вопросы и ответы
 Скачать 0.54 Mb. Скачать 0.54 Mb.
|

|
Задний мозг является частью ромбовидного мозга, формируется из ромбомеров 1, 2 и 3. Дорсальная часть -мозжечок, ножки мозжечка (проводящие пути), которые переплетаются на вентральной стороне в виде массивноговаролиева моста. В глубине стволовой части заднего м 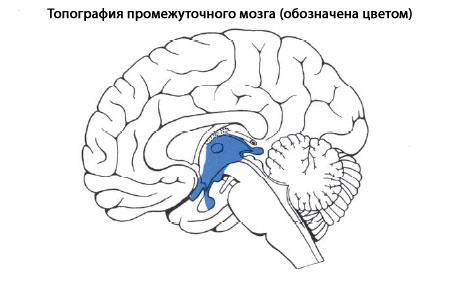 озга лежат ядра черепно-мозговых нервов, проводящие пути, ретикулярная формация, шов. В ядрах шва – центр засыпания. В толще располагается канал - сильвиев водопровод. озга лежат ядра черепно-мозговых нервов, проводящие пути, ретикулярная формация, шов. В ядрах шва – центр засыпания. В толще располагается канал - сильвиев водопровод.Ствол мозга. Строение среднего и промежуточного мозга. Промежуточный мозг, diencephalon, залегает под мозолистым телом и сводом, срастаясь по бокам с полушариями конечного мозга. Соответственно сказанному выше о функции и развитиипереднего мозгавпромежуточном мозгеразличают две основные части: дорсальную (филогенетически более молодую) - thalamencephalon - центр афферентных путей и вентральную (филогенетически более старую) - hypothalamus - высший вегетативный центр. Средний мозг, mesencephalon, развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех холмиков крыши среднего мозга. в среднем мозге человека имеются: подкорковые центры зрения и ядра нервов, иннервирующих мышцы глаза; подкорковые слуховые центры; все восходящие и нисходящие проводящие пути, связывающие кору головного мозга со спинным и идущие транзитно через средний мозг; пучки белого вещества, связывающие средний мозг с другими отделами центральной нервной системы. Соответственно этому средний мозг, являющийся у человека наименьшим и наиболее просто устроенным отделом головного мозга, имеет две основные части: крышу, где располагаются подкорковые центры слуха и зрения, и ножки мозга, где преимущественно проходят проводящие пути. Спинной мозг, лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41-42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, conus medullaris, на уровне II поясничного позвонка. Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков). От conus medullaris отходит книзу так называемая концевая нить, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из п 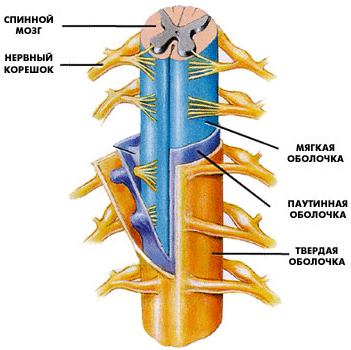 родолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку. родолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку.Спинной мозг на своем протяжении имеет два утолщения, соответствующих корешкам нервов верхней и нижней конечностей: верхнее из них называется шейным утолщением, intumescentia cervicalis, а нижнее - пояснично-крестцовым, intumescentia lumbosacralis. Из этих утолщений более обширно пояснично-крестцовое, но более дифференцировано шейное, что связано с более сложной иннервацией руки как органа труда Наружная, твердая, оболочка спинного мозга отделена от позвоночного столба эпидуральным пространством. Средняя, паутинная, оболочка отделяется от твердой оболочки субдуральным пространством, а от мягкой - подпаутинным. Последнее образует ниже спинного мозга (в области корешков спинно-мозговых нервов - так называемого конского хвоста) терминальный желудочек, заполненный спинно-мозговой жидкостью. Продолговатый мозг и мост к рождению достигают большой степени развития, и отличия их от таковых у взрослого состоят только в степени миелинизации ядер и путей. Их вес у новорожденного составляет 8 г, или 2% массы головного мозга, у взрослого - только 1,6% . По отношению к черепу мост превосходит на 5 - 8 мм спинку седла до 2-летнего возраста, когда устанавливаются соотношения как у взрослого. Строение продолговатого мозга отличается большим ростом медиальной и задней части заднего ядра блуждающего нерва в первые годы жизни. В возрасте около 3 - 4 лет в их клетках появляется пигмент, увеличивающийся в количестве до периода полового созревания. Для новорожденного характерен сегментированный вид задних ядер блуждающего нерва и двойного ядра, которые на продольном срезе имеют более расширенные области, разделенные узкими областями. Дугообразные ядра распространяются более краниально и латерально от ядра оливы и медиально от пирамидных нитей. В части моста четвертого желудочка и его дна - ромбовидной ямки расположена длинная ямка, не пигментированная, называемая locus ceruleus. Пигмент появляется в течение второго года жизни и в возрасте около 10 лет не отличается от пигмента у взрослого. Внутреннее строение моста не представляет никаких особенностей. Пирамидальные пути миелинизированы, корково-мостовые пути еще не миелинизированы. 8. Строение и функции вегетативной нервной системы, ее роль в адаптации организма Вегетативная нервная система выполняет ряд функций: Управляет деятельностью внутренних органов, кровеносных и лимфатических сосудов, осуществляя иннервацию гладкомышечных клеток и железистого эпителия. Регулирует обмен веществ, приспосабливая его уровень к снижению или повышению функции органа. Тем самым осуществляет адаптационно-трофическую функцию, в основе которой лежит транспорт аксоплазмы - процесс непрерывного движения различных веществ от тела нейрона по отросткам в ткани. Одни из них включаются в обмен веществ, другие активируют метаболизм, улучшая трофику ткани. Координирует работу всех внутренних органов, поддерживая постоянство внутренней среды организма. Центры вегетативной нервной системы Центры вегетативной нервной системы разделяют на сегментарные и надсегментарные (высшие вегетативные центры). Сегментарные центры располагаются в нескольких отделах центральной нервной системы, где выделяют 4 очага: Мезенцефалический отдел в среднем мозге - добавочное ядро (Якубовича), nucleus accessorius, и непарное срединное ядро глазодвигательного нерва (III пара). Бульбарный отдел в продолговатом мозге и мосту - верхнее слюноотделительное ядро, nucleus salivatorius superior, промежуточно-лицевого нерва (VII пара), нижнее слюноотделительное ядро, nucleus salivatorius inferior, языкоглоточного нерва (IX пара) и дорсальное ядроблуждающего нерва (X пара), nucleus dorsalis n. vagi. Оба этих отдела объединяются под названием краниального и относятся к парасимпатическим центрам. Тораколюмбальный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 16-ти сегментов спинного мозга (С8, Th1-12, L1-3). Они являются симпатическими центрами. Сакральный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 3-х крестцовых сегментов спинного мозга (S2-4) и относятся к парасимпатическим центрам. Высшие вегетативные центры (надсегментарные) объединяют и регулируют деятельность симпатического и парасимпатического отделов, к ним относятся: Р 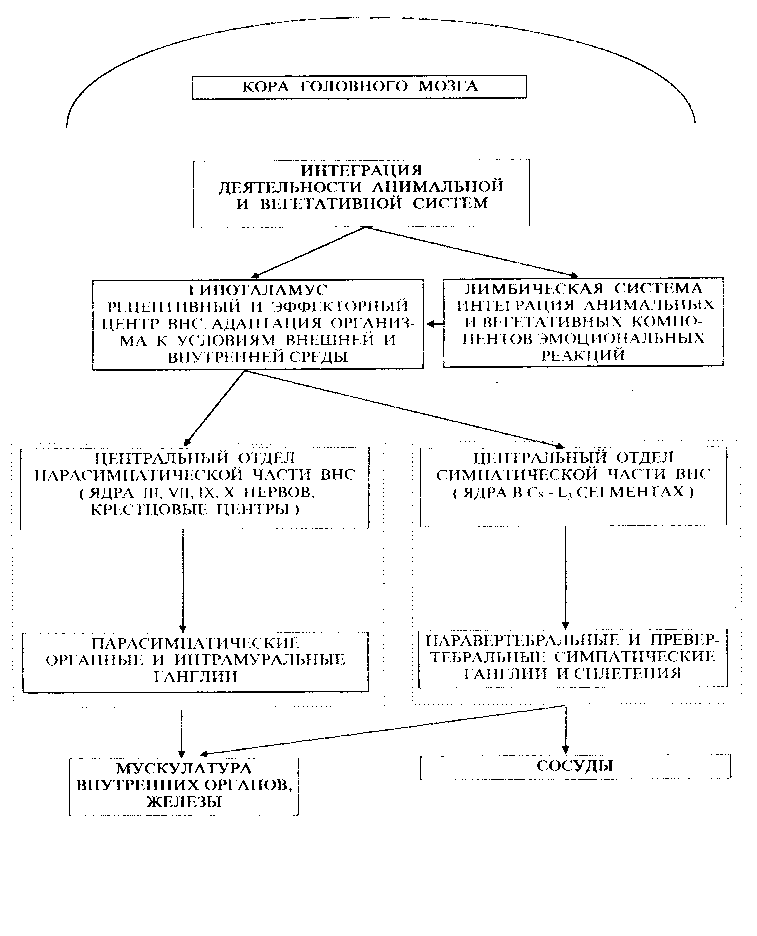 етикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.). Проекция дыхательного центра соответствует средней трети продолговатого мозга, сосудодвигательного центра - нижней части ромбовидной ямки. Нарушение функции ретикулярной формации проявляется вегетативно-сосудистыми расстройствами (кардио-васкулярные, вазомоторные). Кроме того страдают интегративные функции, которые необходимы для формирования целесообразного адаптивного поведения. етикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.). Проекция дыхательного центра соответствует средней трети продолговатого мозга, сосудодвигательного центра - нижней части ромбовидной ямки. Нарушение функции ретикулярной формации проявляется вегетативно-сосудистыми расстройствами (кардио-васкулярные, вазомоторные). Кроме того страдают интегративные функции, которые необходимы для формирования целесообразного адаптивного поведения. Мозжечок, принимая участие в регуляции двигательных актов, одновременно обеспечивает эти анимальные функции адаптационно-трофическими влияниями, которые через соответствующие центры приводят к расширению сосудов интенсивно работающих мышц, повышению уровня трофических процессов в последних. Установлено участие мозжечка в регуляции таких вегетативных функций, как зрачковый рефлекс, трофика кожи (скорость заживления ран), сокращение мышц, поднимающих волосы. Гипоталамус - главный подкорковый центр интеграции вегетативных функций, имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, минерального, водного) и терморегуляции. За счет связей с таламусом он получает разностороннюю информацию о состоянии органов и систем организма, а вместе с гипофизом образует функциональный комплекс - гипоталамо-гипофизарную систему. Гипоталамус в ней выполняет роль своеобразного реле, включающего гипофизарную гормональную цепь в регуляцию различных висцеральных и соматических функций. Особое место занимает лимбическая система обеспечивающая интеграцию вегетативных, соматических и эмоциональных реакций. Полосатое тело имеет ближайшее отношение к безусловнорефлекторной регуляции вегетативных функций. Повреждение или раздражение ядер полосатого тела вызывает изменение кровяного давления, усиление слюно- и слезоотделения, усиление потоотделения. Высшим центром регуляции вегетативных и соматических функций, а также их координации является кора полушарий большого мозга. Непрерывный поток импульсов от органов чувств, сомы и внутренних органов по афферентным путям поступает в кору головного мозга и через эфферентную часть вегетативной нервной системы, главным образом через гипоталамус, кора оказывает соответствующее влияние на функцию внутренних органов, обеспечивая адаптацию организма к меняющимся условиям окружающей и внутренней среды. Примером кортиковисцеральной связи может служить изменение вегетативных реакций под влиянием словесных сигналов (через вторую сигнальную систему). Таким образом, вегетативная нервная система, так же как и вся нервная система, построена по принципу иерархии, подчиненности (рис). 9. Строение, функции и возрастные особенности больших полушарий и коры головного мозга. Локализация функций в коре,функциональная асимметрия полушарий. Кора больших полушарий головного мозга, слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга, развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности, хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см2. Головной мозг новорожденных и дошкольников короче и шире, чем у школьников и взрослых. До 4 лет происходит почти равномерный рост мозга в длину, ширину и высоту, а с 4 до 7 лет особенно интенсивно увеличивается его высота. Отдельные доли мозга растут неравномерно: лобная и теменная доли растут быстрее височной и особенно затылочной. Средний абсолютный вес головного мозга у мальчиков и девочек составляет соответственно (в граммах): у новорожденных – 391 и 388; в 2 года – 1011 и 896; в 3 года – 1080 и 1068; в 5 лет – 1154 и 1168, в 9 – 1270 и 1236. К 7 годам вес мозга соответствует 4/5 веса мозга у взрослых. После 9 лет вес головного мозга прибавляется медленно, к 20 годам он достигает уровня взрослых, а наибольший вес мозг имеет в 20–30 лет. Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними. Пространство между телами и отростками нервных клеток коры заполнено нейроглией и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон. Звездчатые клетки отличаются слабым развитием дендритов и мощным развитием аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением. Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Речь как функция мозга. Кора больших полушарий головного мозга человека содержит три важнейших для речевой функции сенсорных поля: (1) зрительное 2) слуховое 3) соматосенсорное В передней центральной извилине правого и левого полушарий (поля 4 и 6 по Бродману) расположено первичное моторное поле, которое управляет мышцами лица, конечностей и туловища. Именно оно определяет произвольную двигательную активность человека, существенной частью которой является речь и письмо. существуют вторичные сенсорные и моторные поля, расположенные в непосредственной близости к первичным зонам. Взаимодействие перечисленных корковых зон осуществляется как за счет транскортикальных ассоциативных связей, так и корково-таламических и соответствующих таламо-корковых связей. Распределение в правом и левом полушарии мозга такой высокоспециализированной функции человека как речь, глубоко асимметрично. Три взаимосвязанные речевые зоны, расположенные в задней височной области, нижней центральной извилине и в дополнительной моторной коре левого полушария, действуют как единый речевой механизм.. После того, как акустическая информация, заключенная в слове, обрабатывается в слуховой системе и в других "неслуховых" образованиях мозга, она поступает в первичную слуховую кору. Однако, для понимания человеком смысла речи и выработки программы. Для произнесения слова необходимо, чтобы активировалось его представительство в зоне Брока, расположенной в третьей лобной извилине. Активация зоны Брока после понимания смысла речи, благодаря участию зоны Вернике, обеспечивается группой волокон, называемой дугообразным пучком. В зоне Брока сведения, поступившие из зоны Вернике, приводят к возникновению детальной программы артикуляции. Реализация этой программы осуществляется через активацию лицевой проекции моторной коры, управляющей речевой мускулатурой и связанной с зоной Брока короткими волокнами. Если воспринимается письменная речь, то сначала включается первичная зрительная кора. После этого информация о прочитанном слове поступает в угловую извилину, которая связывает зрительную форму данного слова с его акустическим аналогом в зоне Вернике. Дальнейший путь, приводящий к возникновению речевой реакции, такой же, как и при чисто акустическом восприятии. Аналогичный путь восприятия письменной речи и у глухих людей. При повреждении различных участков коры левого полушария и соединяющих эти участки нервных путей возникают нарушения речи - афазии. Формы и проявления афазий различны: это нарушение артикуляции речевых звуков, неспособность к построению осмысленной речи, даже если произнесение звуков не нарушено, это также неспособность понимать устную речь. Корковые отделы левого полушария играют специфическую роль в восприятии, запоминании и воспроизведении речевого материала. Именно эти зоны необходимы для полноценного осуществления речевой функции как единого сенсорного, мыслительного и моторного процесса. Расположенные спереди зоны особенно важны для осуществления экспрессивной (выразительной) речи, расположенные сзади - для восприятия смысла речи. Взаимодействие речевых корковых зон осуществляется не только по горизонтали с помощью корково-корковых связей и взаимодействий, но и по вертикали, через таламические ядра. Другими словами, речевая функция непосредственно связана с функциями различных подкорковых образований мозга. Функциональная асимметрия мозга в связи с механизмами речи проявляется следующим образом. Тональный слух идентичен для обоих полушарий. 10. “Высшая ” и “низшая” нервная деятельность. Роль работ ИМ Сеченова и ИП Павлова в создании учения о В.Н.Д. Павловский метод эксперементального изучения В.Н.Д. ННД направлена во внутреннюю среду организма. Это совокупность нейрофизиологических процессов, обеспечивающих осуществление безусловных рефлексов и инстинктов. ННД – это деятельность СМ и ствола ГМ, обеспечивающая регуляцию деятельности внутренних органов и их взаимосвязь, благодаря чему организм функционирует как единое целое. ВНД направлена на внешнюю среду. Это совокупность нейрофизиологических процессов, обеспечивающих сознательную и подсознательную переработку информации, усвоение информации, приспособительное поведение к окружающей среде и обучение в онтогенезе всем видам деятельности, в том числе целенаправленному поведению в обществе. Это деятельность коры БП и прилегающих к ней подкорковых структур, обеспечивающих взаимосвязь организма с окружающей средой. Важным элементом ВНД является условный рефлекс, т.е. сформированная в процессе онтогенезе реакция организма на раздражитель, ранее индеферентный для этой реакции. ВНД – это аналитико-ситетическая деятельность коры и ближайших подкорковых образований, которая проявляется в способности выделять из окружающей среды ее отдельные элементы и объединять их в комбинации. Важную роль в развитии представлений о ВНД сыграл Сеченов. Он впервые высказал мысль о том, что сознание представляет собой ль субъективное отражение объективной действительности. Он считал ч в основе познания человеком окружающей среды лежит деятельность органов чувств. И.П. Павлову наука обязана всесторонними исследованиями физиологии головного мозга и созданием материалистического учения о высшей нервной деятельности. Для современных представлений о работе мозга решающим явилось открытие И.П. Павловым принципа условнорефлекторной связи —т.е. условного рефлекса. УР – основной и наиболее характерный вид деятельности головного мозга, УР – это основа, на которой строится высшая нервная деятельность, почти все поведение высокоразвитого организма. Метод Павлова позволил исследовать образование условных и безусловных рефлексов. Павлов обратил внимание, что желудочный сок у собаки выделяется не только при виде пищи, но и когда животное слышит шум шагов человека, который ее приносит. Так ученый начал исследовать и процессы жизнедеятельности головного мозга. Причем реакции подобного рода могли вырабатываться у животных не только в ответ на шум шагов, но и на включение света, звон колокольчика, различные запахи и т. д. Физиолог разделил все возможные ответы организма на две категории. Врожденные реакции он назвал безусловными, а те, что были приобретены в процессе жизнедеятельности – условными. К первой категории относится защита от врагов, поиски пищи, а также достаточно сложные действия – например, строительство гнезда. Безусловные рефлексы присущи каждому живому организму с рождения. А выполнение тех команд, которые животное получает от дрессировщика, относится к категории условных рефлексов. 11. Виды рефлексов, отличие условных рефлексов от безусловных. Приспособительное значение условных рефлексов. Механизм выработки условного рефлекса. Условные и безусловные рефлексы имеют единую материальную основу – нервные процессы. В поведенческих актах условные и безусловные рефлексы представляют собой своеобразный сплав, единство врожденного и приобретенного. У взрослых животных «чистые» безусловные рефлексы практически не обнаруживаются. Все они как бы обрастают условными рефлексами. Различия между этими двумя группами рефлексов носят относительный характер, но они важны для понимания механизма образования условнорефлекторных связей. Выработка условных рефлексов Рефлексы названы условными потому, что возникают (и в природе, и в лаборатории) лишь при определенных условиях. Одними из них являются: Неоднократное сочетание ранее индифферентного условного раздражителя с действием подкрепляющего безусловного (или ранее хорошо выработанного условного) раздражителя; Некоторое опережение во времени влияния индифферентного агента действию подкрепляющего раздражителя; Бодрствующее состояние организма; Отсутствие других видов активной деятельности и посторонних раздражителей; Достаточная степень возбудимости животного к действию безусловного подкрепляющего раздражителя; Надпороговая интенсивность условного раздражителя. В лабораторных условиях формирование классического условного рефлекса начинается с погашения ориентировочной реакции животного на раздражитель, который в дальнейшем должен быть условным сигналом. Так, если перед собакой зажечь лампочку, то вначале у нее возникает ориентировочный рефлекс (поворот головы, туловища, движение глаз на свет). При повторном зажигании лампочки ориентировочная реакция уменьшается, а затем угасает. Свет лампочки становится индифферентным раздражителем. В дальнейшем этот раздражитель действует изолированно в течение 5 – 10 сек., после чего к нему присоединяется безусловный раздражитель (корм). Совместное действие раздражителей продолжается в течение периода еды. Такое сочетание условного сигнала и безусловного раздражителя повторяют несколько раз (за один опыт 8 – 10 сочетаний). Через несколько сеансов зажигание лампочки будет вызывать отделение слюны без пищевого подкрепления, что свидетельствует о выработке слюноотделительного условного рефлекса. Аналогичным образом вырабатывается и двигательно-оборонительный условный рефлекс, когда индифферентный раздражитель сочетается с болевым раздражением конечности животного. Показателем образования рефлекса служит сгибание лапы (устранение от опасности) при действии одного условного раздражителя. Несколько сложнее задача у животного при выработке оперативного (инструментального) условного рефлекса. В отличие от классических условных рефлексов подкрепление здесь дается лишь после определенной двигательной реакции животного в ответ на условный раздражитель (нажатие на рычаг, дерганье зубами кольца, открывание дверцы, пробежка по лабиринту). Сначала эта реакция животных может проявиться случайно, затем становится закономерной.
|