Курсовая_брачное поведение животных. Функции брачного поведения
 Скачать 318 Kb. Скачать 318 Kb.
|

СодержаниеВведение 2 Функции брачного поведения. 4 Системы брачных отношений (моногамия, полигамия, полигиния, полиандрия, полибрахигиния, промискуитет). 4 Сезонность и синхронизация 7 Соблазнение и умиротворение. 133 Репродуктивная изоляция. 200 Формы ухаживания у различных видов: Членистоногие. 23 Рыбы 24 Амфибии и рептилии. 25 Птицы 25 Млекопитающие 206 Заключение 278 Список использованной литературы 29 Введение Многие животные, особенно морские, обеспечивают оплодотворение яйцеклеток таким простым способом, что говорить о каком-то особом брачном поведении затруднительно. Например, птицы просто извергают в воду огромные количества своих спермиев в определенное время года, каждая особь окружается целым их облаком. По-видимому, яйцеклетки просто не могут избежать оплодотворения. Однако даже здесь различим важный поведенческий аспект: оплодотворения не произойдет, если различные особи устриц не образуют свои спермин и яйцеклетки одновременно. Значит, необходима определенная синхронизация. У многих высших животных, особенно сухопутных, оплодотворение требует спаривания, или копуляции. Для этого необходима не только синхронизация, но и физический контакт особей. Его большинство животных избегают. Избегание — адаптация, частично обеспечивающая защиту от хищников: прикосновение к телу обычно означает, что тебя поймали. Кроме того, во время самого спаривания животные (особенно самки) находятся в беспомощном положении. Другими словами, брачное поведение нередко включает в себя подавление защитных реакций. Поскольку самка некоторое время вынашивает яйца, часто даже после их оплодотворения, и поскольку у многих видов именно она играет главную роль в выкармливании и защите потомства, она составляет наиболее ценную часть «видового капитала». Кроме того, один самец часто способен оплодотворить несколько самок, и по этой причине отдельные самцы также менее ценны биологически. Таким образом, неудивительно, что самка больше, чем самец, нуждается в «соблазнении» партнером. Вероятно, поэтому ухаживание так часто входит в задачи именно самца. Иногда приходится «соблазнять» и его, но уже по другой причине. Самцы большинства видов в сезон размножения крайне агрессивны, и пока самкам не удастся их «умиротворить», они вместо ухаживания могут на них напасть. Далее, помимо синхронизации, означающей координацию спаривания во времени, требуется тесная пространственная координация: самцы и самки должны друг с другом встретиться, во время спаривания привести в соприкосновение свои половые органы, и, наконец, спермий должен найти яйцеклетку. Эта ориентация в пространстве также составляет часть брачного поведения. Наконец, очень важно избежать спаривания с представителями других видов. Поскольку гены и крайне сложные запускаемые ими ростовые процессы у каждого вида особые, скрещивание особей разных видов приведет к сочетанию несовместимых генов, а это легко может нарушить тонко сбалансированный ход развития потомства. В результате такое спаривание часто приводит к нежизнеспособным оплодотворенным яйцеклеткам, погибающим в самом начале развития. В менее тяжелых случаях гибриды выживают, но становятся слабыми или бесплодными. Необходимость избегать межвидового скрещивания привела к формированию видоспецифичных особенностей брачного поведения, так что каждая особь без труда «распознает» партнера собственного вида. Таким образом, кроме собственно осеменения к функциям брачного поведения относятся синхронизация, ухаживание, пространственная ориентация и репродуктивная изоляция. В данной работе мы рассмотрим, как выполняются перечисленные функции, какую роль играет социальное поведение, и как оно приводит к нужным результатам. Функции брачного поведения.Функция ухаживания состоит в том, чтобы обеспечить обнаружение и опознание потенциальных партнеров, принадлежащих к соответствующему полу и виду, и синхронизировать их репродуктивную активность. Иначе говоря, функция ухаживания состоит в том, чтобы свести вместе двух животных разного пола в условиях, обеспечивающих большую вероятность успешного спаривания. Прежде всего необходимо, чтобы животное могло найти потенциального брачного партнера. Ухаживание нередко представляет собой сложную последовательность взаимодействующих сигналов, которые должны привести к спариванию данной особи с подходящим партнером. Для успешного размножения большое значение имеют сроки; при спаривании как самец, так и самка должны находиться в надлежащем физиологическом состоянии. Это достигается синхронизацией циклов, которую обеспечивает взаимодействие внешних стимулов и поведения самих партнеров. Представители классической этологии указывали, что многие формы ухаживания содержат элементы конфликта, нередко по той причине, что первоначальная реакция особи на появившуюся поблизости другую особь может быть агрессивной. Системы брачных отношений (моногамия, полигамия, полигиния, полиандрия, полибрахигиния, промискуитет). Популяции животных характеризуются определенными системами брачных отношений. Подавляющее большинство видов насекомоядных и хищных птиц, а также хищных млекопитающих моногамны, т. е. брачную пару образует один самец и одна самка. В подобных случаях обычно оба родителя принимают участие в воспитании потомства. Полигамия—это такая система брачных отношений, при которой одна особь вступает в брачную связь одновременно с более чем одним представителем противоположного пола. Существуют два типа полигамии, в зависимости от того, какой пол поддерживает множественные связи. У некоторых птиц, например у желтоголового трупиала, один самец имеет одновременно брачные связи с двумя или большим числом самок (полигиния). Значительно реже встречается полиандрия, при которой одна самка поддерживает брачные отношения с более чем одним самцом. Полиандрия, по-видимому, наблюдается иногда у птиц... При полибрахигинии один самец вступает в кратковременную брачную связь с несколькими самками поочередно; в типичном случае такая связь устанавливается только на время, необходимое для копуляции и осеменения. Полибрахигиния встречается у ряда птиц (в том числе у некоторых скворцов, колибри и тетеревиных) и млекопитающих (многих ластоногих и некоторых копытных). Наконец, идеальная структура брачных отношений (или, может быть, правильнее сказать отсутствие таковых) – это промискуитет, когда любая особь имеет равную вероятность скрещивания с любой другой особью. Истинный промискуитет, приводящий к панмиктической популяции, в высшей степени маловероятен и, по-видимому, не существует. Может быть, отчасти к нему приближаются некоторые многощетинковые черви и морские лилии, выпускающие свои гаметы просто в море, или наземные ветроопыляемые растения, т. е. такие организмы, гаметы которых переносятся водными течениями или воздушными потоками. Однако даже у этих прикрепленных организмов, по-видимому, встречаются различные формы химической дискриминации гамет и; соответственно — предпочтения при скрещивании. Тот компонент полового отбора, который связан с отношениями представителей разных полов, называется эпигамным отбором.Его часто определяют как «возрастание репродуктивного преимущества тех генотипов, которые обеспечивают наиболее сильные гетеросексуальные стимулы», или также как «борьбу полов». Эпигамный отбор оперирует предпочтением в выборе брачного партнера. Первостепенное значение имеет тот факт, что особенности, максимизирующие приспособленность самца, не обязательно совпадают с таковыми особенностями самки, и соответственно наоборот. Например, у большинства позвоночных животных самец обычно может оставить больше генов в условиях полигинии или полибрахигинии, тогда как самка, по-видимому, достигнет максимального репродуктивного успеха при моногамии или полиандрии. Поскольку сперматозоиды мелки и их образование требует незначительных энергетических затрат, они образуются в больших количествах, а так как вклад самцов позвоночных в каждый акт размножения сравнительно мал, они могут спариваться часто и достаточно неразборчиво (иными словами, самцы тяготеют к промискуитету). Самки позвоночных, наоборот, обычно вносят гораздо больший вклад в каждый акт размножения, так как образование яиц или детенышей требует больших энергетических затрат. Из-за того что самки гораздо сильнее связаны с каждым актом размножения, они склонны предъявлять значительно более строгие требования к брачному партнеру, чем самцы, и гораздо более разборчивы. Отказываясь спариваться с самцами, которым свойствен промискуитет, самки позвоночных могут иногда «заставить» самцов стать моногамными и внести свою долю в воспитание потомства. Фактически полигиния – это такой исход борьбы полов, при которой выигрывают самцы (патриархат), а полиандрия – при которой выигрывают самки (матриархат). Моногамия представляет собой компромисс между этими двумя крайними вариантами. При моногамной системе брачных отношений самец должен быть уверен в том, что потомство, которое он выращивает, его, так как в противном случае он может тратить энергию на чужого самца (заметьте, что для самок такой проблемы не возникает). Неудивительно, что моногамные самцы ревниво охраняют своих самок от копуляций с посторонними самцами. Вместе с тем самки, связанные моногамными отношениями, очень уязвимы дезертирством самцов в случае начавшегося размножения. Сезонность и синхронизация.Одна из главных особенностей цикла размножения у многих видов – его сезонность, т. е. приуроченность к определенным временам года. Многие животные размножаются в определенное время года, с тем чтобы рождение потомства приходилось на период, оптимальный в отношении ресурсов среды. Весенний пик репродуктивной активности у птиц послужил источником вдохновения для многих писателей и поэтов. Другие животные, однако, размножаются в иное время года. Многие виды оленей и лоси размножаются осенью, волки и койоты – в середине зимы, некоторые тюлени и морские львы – в конце весны и в начале лета. В пределах данного вида сезоны размножения часто варьируют в зависимости от географической широты. Общим знаменателем для всех этих различных сезонов размножения служит время рождения детенышей. Хотя у разных животных сроки беременности различны, большинство из них рождает детенышей в конце весны и начале лета. По-видимому, главное преимущество сезонности – то, что она позволяет приурочить появление потомства к хорошей погоде и наибольшему обилию пищевых ресурсов. Брачные сезоны и периоды беременности, по-видимому, согласованы между собой таким образом, что у большинства видов детеныши рождаются в конце весны и в начале лета. Факторы, с которыми непосредственно связано начало репродуктивной активности, у разных видов различны. Такие условия, как температура, количество осадков, развитие растительности и длина дня, изменяются в зависимости от времени года, и все они могут влиять на репродуктивную активность у тех или иных животных. По-видимому, начало размножения у многих видов определяется длиной дня, поскольку во многих местообитаниях этот фактор наиболее надежным образом коррелирован со сменой времен года. Так, например, если содержать дальневосточного перепела в условиях короткого дня (8 ч света и 16 ч темноты), то как у самцов, так и у самок репродуктивные органы уменьшаются и половые реакции исчезают. Надлежащее изменение светового режима (16 ч света и 8 ч темноты) приводит к восстановлению как морфологии, так и поведения. Хронология репродуктивного поведения у устриц (Ostreaedulis) регулируется довольно неожиданным внешним фактором, поэтому, строго говоря, не относится к «социологической» проблематике. Однако не исключено, что полезно будет рассмотреть ее здесь в качестве примера того, как действие внешних факторов может, так сказать, «имитировать» социальное сотрудничество. Примерно через восемь дней после икрометания устриц начинается «роение» их личинок. Они очень недолго плавают, а затем оседают на твердый субстрат. В илистых эстуариях Шельды в Голландии устрицеводы увеличивают свои «стада», укладывая на морское дно черепицу, служащую личинкам устриц искусственным субстратом. Этого нельзя делать задолго до массового появления личинок, поскольку куски черепицы будут быстро заняты другими организмами. Таким образом, от зоолога требуется определить, если это возможно, когда произойдет «роение». Прогноз, основанный на многолетних исследованиях, звучит удивительно: «Основной максимум появления личинок следует ожидать ежегодно между 26 июня и 10 июля, примерно через 10 дней после новолуния или полнолуния» (рис. 13). Это кажется мистикой, но является чистой правдой. Поскольку «роение» происходит через восемь дней после икрометания, значит, самого икрометания следует ожидать через два дня после новолуния или полнолуния. Отсюда выводится фактор, определяющий время размножения, прилив. Икрометание происходит при сизигийном приливе. Как он влияет на устриц, еще неизвестно, не исключено, что это связано с давлением воды, колебания которого максимальны именно при сизигийном приливе. В это же время больше всего колебания освещенности на дне моря: чем не фактор?  Рис.1. Массовое появление личинок устриц в течение 74 последовательных дней в июне, июле и августе. Заметна корреляция с фазами Луны. Поскольку устрицы не мечут икру при каждом сизигийном приливе, должен быть и другой фактор, подготавливающий их именно к июньской реакции на такой прилив; природа его еще неизвестна. Он действует не слишком точно, поскольку, хотя максимум икрометания приходится на период 18 июня — 2 июля, меньшие по высоте пики наблюдаются и при других сизигийных приливах. Для устриц этот фактор неизвестен, но в случае других животных мы кое-что о нем знаем. Время размножения связано с приливами не только у устриц, но и у некоторых других обитателей моря. Среди них знаменитый тихоокеанский червь палоло и еще несколько видов червей и моллюсков. Временной фактор у высших животных — более сложная проблема. Кое-что известно о рыбах, птицах и млекопитающих северной умеренной зоны. Размножение у большинства из них начинается весной. Первая фаза — миграция к местам размножения. Она происходит у всех особей вида примерно одновременно, хотя между прибытием на место первой и последней из них иногда проходят недели. Эта грубая синхронизация обусловлена опять-таки не социальным поведением, а реакцией на внешние факторы. В данном случае главный из них — удлинение светового дня в конце зимы. Известно, что различные млекопитающие, птицы и рыбы реагируют на искусственное удлинение долготы дня. В результате их гипофиз начинает секретировать гормон, который в свою очередь влияет на рост половых желез. Они в свою очередь секретируют половые гормоны, а действие последних на центральную нервную систему запускает первую поведенческую реакцию, связанную с размножением,— миграцию. Дополнительное влияние часто оказывает повышение температуры среды. Как уже говорилось, эта синхронизация не очень точная. Отдельные особи реагируют на удлинение светового дня неодинаково интенсивно, причем могут наблюдаться значительные различия даже между самцом и самкой одной пары. В случае голубей и некоторых других животных было замечено, что, если самец реагирует раньше самки, его настойчивые ухаживания могут ускорить гормональное развитие последней. Это обнаружилось следующим образом. Когда самца и самку держали раздельно в соседних клетках так., чтобы они могли видеть и даже касаться друг друга, но не копулировать, непрерывное ухаживание голубя в конце концов индуцировало голубку снести яйца. Естественно, они оказались нежизнеспособными. В неволе бывает, что в отсутствие самцов пару образуют две голубки, одна из которых демонстрирует поведенческие особенности, обычно присущие самцу. Хотя их репродуктивные ритмы сначала могут быть разными, в результате обе несут яйца одновременно. Действия в отношении друг друга как-то стимулируют синхронизацию, причем не только поведенческую, но и связанную с развитием яиц в яичниках. Возможно, этот эффект встречается и у других видов. Как полагал Дарлинг, он проявляется и при массовом ухаживании в парах птиц, размножающихся внутри колонии. Однако необходимо дальнейшее повышение точности синхронизации. У всех копулирующих (да и у многих других) видов сотрудничество между самцом и самкой должно разворачиваться в соответствии с точным «расписанием», и без тонко настроенной синхронизации оплодотворение невозможно. Только у очень немногих животных самец способен заставить самку спариваться с ним насильно. Значит, чаще всего какая-то форма очень точной синхронизации должна осуществляться на уровне долей секунды. Это обеспечивается своего рода сигнальной системой. В качестве примера рассмотрим спаривание трехиглой колюшки. Каждая реакция действует как сигнал, запускающий следующую реакцию у партнера. Так, зигзагообразный танец самца заставляет приближаться к нему самку, в ответ на это он ведет ее к гнезду, она реагирует, следуя за ним, и т. п. Это легко показать с помощью моделей или макетов. Когда самцу на его гнездовой территории показывают очень грубое подобие самки с полным икры брюшком (рис.2), он приближается к нему и исполняет зигзагообразный танец. Как только модель разворачивается в его направлении и «плывет» навстречу, он начинает вести ее к гнезду. 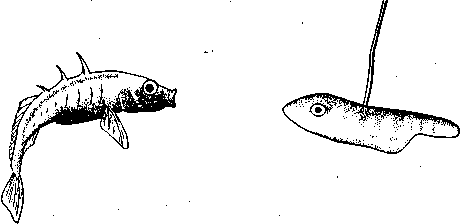 Рис.2. Самец трехиглой колюшки ухаживает за грубым макетом самки. Самку с икрой можно аналогичным образом заставить реагировать на модель самца. И тут будет достаточно самой грубой подделки, лишь бы нижняя часть макета была красной, а глаза ярко-голубыми. Остальные детали не важны. Если такую модель перемещать вокруг самки, очень поверхностно имитируя зигзагообразный танец, самка развернется и приблизится к ней. Если теперь модель «поплывет» прочь, самка последует за ней и можно даже заставить ее попытаться проникнуть в любую дырку на дне аквариума, продемонстрировав моделью самца «вход в гнездо» (рис.3). В самом гнезде нет необходимости, движения модели — достаточный стимул для реакции самки. В этих случаях рыба реагирует не только на перемещения партнера, но и на некоторые черты его формы и окраски. Если у макета самки «брюхо» не вздуто, танец самца, если и будет стимулироваться, то очень слабо. Если у модели самца нижняя часть не красная, самка не проявит к ней интереса. С другой стороны, все прочие детали практически не важны, поэтому легче запустить брачное поведение самца, используя очень грубую, но «икряную» подделку, чем живую, но не готовую к размножению самку. Однако сами по себе ни вздутое, ни красное брюхо, сколько бы их ни предъявлять, не вызывают синхронной реакции у соответствующего пола. Она возникает только в ответ на определенное движение, которое таким образом и обусловливает точную синхронизацию событий. 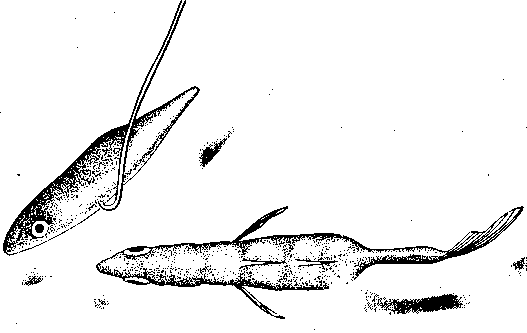 Рис.3. Самка трехиглой колюшки следует за макетом самца, «показывающим вход в гнездо». Вид сверху. Брачное поведение трехиглой колюшки — это сложный ряд таких сочетаний сигнал — ответ, приводящих в конечном итоге к оплодотворению самцом икринок сразу же после их выметывания самкой. Наблюдать подобные действия и проводить описанные выше опыты с моделями совсем не трудно. Эта рыба легко размножается в аквариуме емкостью около 20 л. На дно следует насыпать песок и поместить побольше зеленой растительности, включая нитчатые зеленые водоросли. С такими сигнальными движениями, непосредственно обеспечивающими высокую точность синхронизации, связано брачное поведение многих видов. Соблазнение и умиротворение.Даже когда животное сексуально возбуждено, оно не всегда сразу же реагирует на ухаживание партнера. Преодоление нежелания самки спариться может потребовать много времени. Так, Зигзагообразный танец колюшки не всегда немедленно вызывает ее реакцию. Самка может нерешительно приблизиться к самцу и остановиться, когда он попытается увести ее к гнезду. В этом случае самец возвращается и снова исполняет зигзагообразной танец. После нескольких его повторов самка в конце концов иногда (но не всегда) соглашается следовать за ним и входить в гнездо». Сходное повторение сигналов необходимо, когда самка уже в гнезде. Чтобы она выметала икру, самец должен ее как следует «потормошить». Если убрать его сразу, после того как самка вошла в гнездо, икрометания не произойдет. Однако, если мягко дотрагиваться до нее стеклянной палочкой, имитируя совершаемое самцом тормошение, икра будет отметана так же легко, как и в случае стимуляции, исходящей от него самого. И самец, и палочка должны дотрагиваться до самки многократно. 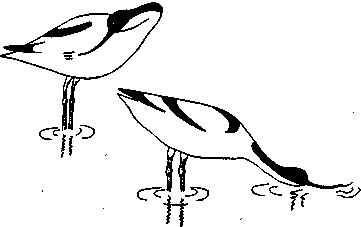 Рис.4. Предкопуляционная демонстрация у шилоклювки. У многих видов такое повторение сигналов — правило. Например, копуляции шилоклювок предшествуют любопытные ужимки: самец и самка стоят рядом и как-то торопливо, «нервно» чистят свои перья. Спустя некоторое время самка прекращает охорашиваться и вытягивается горизонтально (рис. 4). Это сигнал, что она готова к спариванию. Только теперь самец взлетает на нее, и происходит копуляция. Иногда он реагирует не сразу, а через некоторое время. 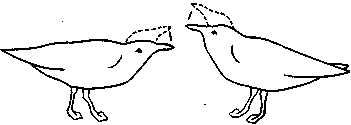 Рис.5. Предкопуляционная демонстрация у серебристой чайки. У серебристой чайки копуляции предшествует аналогичное вступление. И самец, и самка неоднократно вскидывают вверх головы, каждый раз издавая при этом мягкие, мелодичные призывы (рис. 5). В данном случае инициативу берет на себя самец: после ряда таких сигналов он неожиданно вскакивает на самку и спаривается с ней. 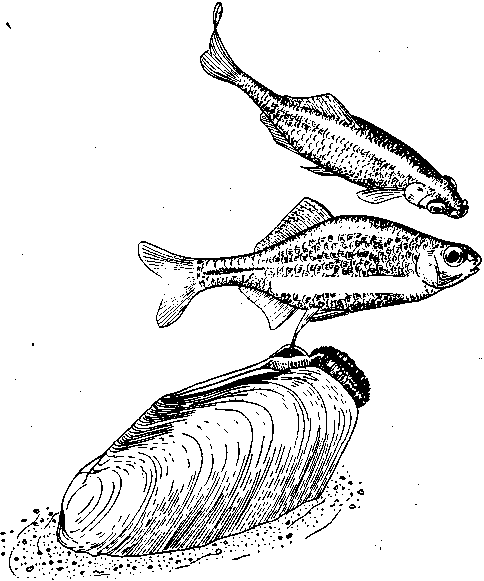 Рис.6. Самец горчака ухаживает за самкой во время икрометания. Иногда «соблазнение» выполняет другую функцию. У многих птиц, да и у других животных самцы в сезон размножения становятся очень агрессивными. В самом деле, большинство сражений в природе происходит весной между соперничающими самцами. В этих драках вся причина. Поскольку нападения всегда направлены против самца-соперника, самка должна отличаться от него, чтобы не быть атакованной. У таких видов, как зяблик, горихвостка или фазаны, этой цели частично служат различия в оперении. Однако у многих других видов, например у крапивника, окраска обоих партнеров почти одинакова, поэтому самке, чтобы подавить агрессивность самца, приходится особым образом себя вести. Значит, задача такого «самочьего ухаживания» — постараться не спровоцировать нападения. Если самец-чужак либо спасается от демонстрирующего агрессивность хозяина территории (что немедленно индуцирует преследование), либо хорохорится и угрожает в ответ (что также вызывает демонстрацию агрессивности), самка ничего такого не делает. У рыбы горчака (Rhodeusamarus) в ответ на первую атаку она либо спокойно отступает, либо избегает столкновения, проплывая под самцом. Видимо, после этого самец уже не способен напасть на нее, спустя некоторое время полностью утрачивает агрессивность и начинает ухаживание (рис. 6). Сходное ненавязчивое умиротворение можно наблюдать у многих цихловых рыб. В других случаях самка демонстрирует инфантильное поведение, т. е. прибегает к такому же способу умиротворения, что и потомство, возможно, стимулируя тем самым у самца родительские побуждения. Именно поэтому у очень многих видов самцы при ухаживании кормят самок; уже говорилось, что такое свойственно серебристой чайке. Однако у некоторых видов умиротворяющая поза при ухаживании отличается от используемой ювенильной особью. Самка (а иногда и оба партнера) демонстрирует поведение, которое во многих смыслах является противоположным угрожающему. Когда, например, обыкновенные чайки (Lamsridubundus) встречаются в сезон размножения, они применяют «впередсмотрящую демонстрацию», опуская голову и направляя клюв друг на друга (вкладка 2, вверху). Эта угрожающая поза подчеркивается коричневой окраской их «лица» вокруг клюва, являющегося серьезным оружием. В то же время половые партнеры демонстрируют свои лучшие намерения с помощью, так сказать, «отмашки головой», т. е. вытягивая вверх шеи, резким рывком отворачиваются друг от друга. При этом, поскольку оба пола достаточно агрессивны, самец умиротворяет самку, а самка — самца. У некоторых тенетных пауков самец посещает самку на ее паутине. В данном случае он должен ее умиротворить, чтобы она не приняла его за добычу. Пространственное управление брачным поведением — еще одна важная функция ухаживания. Наиболее очевидный ее пример — привлечение. Многие певчие птицы, например соловьи, проводят зиму вдали от гнездовых участков. Самцы, как уже говорилось выше, возвращаются с юга гораздо раньше самок. Как же самки их находят? По песне. Многие птицы привлекают противоположный пол какими-нибудь громкими звуками. У соловья эти призывы приятны человеку, поэтому он называет их песней. А вот весенние крики самца серой цапли (Ardeadrierea) очень грубы и человеческому уху отвратительны. Однако самку этого вида они привлекают и выполняют точно такую же функцию, что и соловьиное пение. К той же категории звуков относятся треск козодоя, барабанная дробь дятла и кваканье жаб. Пение многих птиц так точно настроено на привлекающую функцию, что достигает максимальной громкости, когда самец одинок, и прекращается, как только у него появляется самка. Здесь перед нами опять же столкновение различных интересов. Пение служит для привлечения самок (и, как будет показано ниже, для отпугивания самцов-соперников), но опасно для самца, поскольку выдает его хищникам. Природа, как всегда, пошла на компромисс: песня звучит до тех пор, пока действительно нужна или по крайней мере пока ее преимущества перевешивают недостатки. Поскольку многие животные глухи (только позвоночные и некоторые другие группы составляют исключение), звуковые демонстрации встречаются относительно редко. Они хорошо развиты у птиц, жаб и лягушек, а также у ряда насекомых, включая сверчков и кузнечиков. У этих животных сформировались особые 'органы, предназначенные исключительно для издавания звуков.  Рис.7. Saturniapyri. У самца на антеннах сильно развиты хемо-рецепторные органы. Другие группы используют для привлечения противоположного пола запах. Крайнее выражение этой способности наблюдается у бабочек. Некоторые исследования проводились на семействе мешочниц (Psychidae), которое и будет рассмотрено в качестве примера. Самка здесь утратила способность к полету и практически бескрыла. Вскоре после выхода из куколки она покидает трубчатый чехол из листьев, в котором проходило ее предшествующее развитие, и подвешивается к нему снизу. Самцы могут летать. Выйдя из куколки, они оставляют чехол и отправляются на поиски самок. Их путь направляется запахом, испускаемым девственной самкой. Такой способ привлечения высоко развит и у многих других бабочек, включая виды павлиноглазок (Saturnia, рис. 7) и коконопрядов (Lasiocampa), причем самец часто способен отыскать самку на весьма значительном расстоянии, поскольку его органы обоняния, расположенные на перистых антеннах, высокочувствительны. Гусениц этих видов нетрудно обнаружить, собрать, дать им окуклиться и выйти из куколки, чтобы затем пронаблюдать, как в дом залетают посторонние самцы, привлеченные запахом девственных самок. У многих видов важно зрительное привлечение. Оно прекрасно развито у трехиглой колюшки. Самец приобретает особенно яркую брачную окраску, окончив строить гнездо. Его красное брюхо становится еще более насыщенным по цвету, а темный оттенок спины сменяется флуоресцирующим голубоватым. Одновременно меняется и его поведение. Если во время постройки гнезда самец движется мягко, избегая рывков, то теперь он мечется по своей территории, что вместе с броским внешним видом делает его заметным издалека.  Рис.8. Чибис в полете. 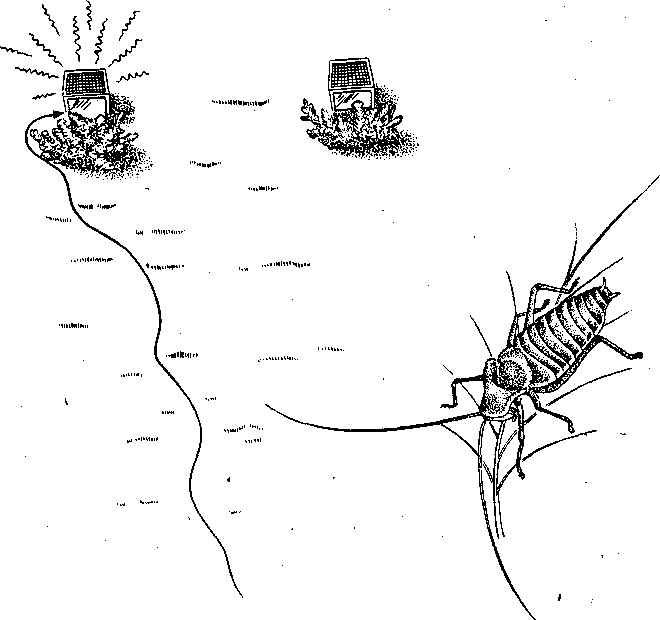 Рис.9. Эксперимент, демонстрирующий функцию песни у кузнечикового Ephippiger. Многие птицы дополняют звуковое привлечение зрительными демонстрациями. Это наиболее развито у видов обширных равнинных пространств. Так, в частности, специализированы многие кулики арктической тундры и европейских болот (рис. 8). Причем опять же часто наблюдается сочетание броской окраски и особых движений. Чибис, большой веретенник, чернозобик — лишь некоторые типичные примеры. У других видов специализированы только привлекающие движения, а внешность неприметная, к ним относятся наиболее уязвимые для хищников певчие птицы типа коньков и жаворонков. Встречается и чисто цветовая специализация. У самца турухтана (Philomachuspugnax) нет особого брачного полета или призывной песни, но он может рассчитывать на окраску своего горлового оперения. Впрочем, у него развилось другое сигнальное движение: время от времени самцы на «току» поднимают крылья, светлая нижняя сторона которых делает их очень заметными. Такой демонстрацией они в основном реагируют на пролетающих вдали самок; по-видимому, она их привлекает. Эти токующие птицы применяют и так называемый «принцип клумбы»: когда они собираются вместе, их индивидуальные расцветки сочетаются друг с другом и образуется крупное пестрое пятно, напоминающее клумбу. Лишь в немногих из этих случаев привлекающее действие доказано экспериментально. Так, ясно, что самки колюшки реагируют на красный цвет брюха самца, модели с другой окраской на них не действуют. Роль звука изящно продемонстрирована для различных кузнечиковых. Один из таких опытов изображен на рис. 9. В клетке, спрятанной в зарослях вереска, держали стрекочущих самцов эфиппигера (Ephippiger); в другой клетке находилось столько же самцов, которые не могли издавать звуков, потому что их стридуляционные органы были склеены экспериментатором. Это простейшая операция, позволяющая таким бескрылым формам свободно осуществлять любую другую активность. На расстоянии 25 м от клеток были выпущены готовые к спариванию самки. Они безошибочно устремились к клетке со стрекочущими самцами. Эксперименты такого типа подтверждают выводы, сделанные выше относительно привлекающего воздействия различных типов демонстраций, хотя, конечно, у экспериментаторов еще много работы. Ориентационная функция ухаживания не ограничивается привлечением. Во время самого спаривания партнеры должны привести свои копуляционные аппараты в соприкосновение, что опять же требует направляющих действий. Это особенно хорошо заметно на примере многих насекомых, самцы которых обладают сложной системой класперов, требующих точной подгонки к соответствующим «обратным» частям полового аппарата самки. Однако и у проще устроенных животных типа птиц такая проблема тоже существует: самец не может привести свою клоаку в соприкосновение с клоакой самки, не реагируя предварительно на поступающие от последней ориентационные стимулы. Впрочем, об этих поведенческих механизмах известно слишком мало. Репродуктивная изоляция.Межвидовая гибридизация происходит в природе исключительно редко. Это лишь отчасти объясняется неодинаковыми требованиями видов к местообитанию. Действительно, близкородственные формы, размножающиеся в отдаленных друг от друга географических областях или живущие в одной области, но размножающиеся в различных биотопах, предохранены от скрещивания пространственной изоляцией. Однако, даже если бы ее не существовало, обычным способом им не скреститься. Это обусловлено тем, что многочисленные сигналы, служащие для привлечения, соблазнения, умиротворения и синхронизации, у каждого вида совершенно своеобразны. Видоспецифична и соответствующая реакция. У каждого животного существует врожденная тенденция как подавать особые, свойственные только своему виду сигналы, так и реагировать исключительно на них. Однако в природе часто можно заметить половую реакцию на другой вид. Самцы бабочки сатира, которых я изучал несколько сезонов, начинают ухаживание, преследуя в полете партнера. Это брачное преследование вызывается не только самками сатира, но и другими бабочками, жуками, мухами, мелкими птицами, опадающими листьями и даже их собственной тенью на земле. Как же получается, что они никогда не спариваются с животными других видов? Сходные наблюдения, приводящие к такому же вопросу, можно провести над птицами, рыбами и многими другими животными. Ответ, по-видимому, стоит искать в цепном характере действий, ведущих к образованию пар и оплодотворению. Когда самка сатира готова к спариванию, она особым образом реагирует на преследование партнера — приземляется. Все прочие виды, как правило, поступают наоборот: потревоженные преследующим их самцом, они улетают с максимально возможной скоростью, так что тот в конце концов от них отстает. Правда, близкородственные виды могут случайно отреагировать на него, но никогда не наблюдалось, чтобы это привело к спариванию. У колюшек в принципе сходное поведение. Самец может среагировать на мелкого линя, привлеченного на его территорию зигзагообразным танцем. Однако для продолжения брачного поведения необходимо, чтобы партнер к нему подплыл. Даже если линь случайно так и поступит, он должен затем следовать за самцом к гнезду, вползать туда и выметывать икру до эякуляции колюшкой спермы. Другими словами, самка другого вида должна продемонстрировать правильную серию реакций на полную последовательность действий по ухаживанию, включая заключительное «тормошение». А это настолько невероятно, что никогда не наблюдалось. Знаковых стимулов каждой отдельной реакции цепочки бывает и недостаточно для предотвращения реакции на другой вид, но, поскольку все эти реакции запускаются различными стимулами, их совокупности, как правило, хватает, чтобы межвидового спаривания не произошло. Это очевидно в случае видов с «взаимным» ухаживанием, так как здесь каждый пол демонстрирует серию специфических брачных действий. Но даже у таких видов, как бабочка сатир, у которых самка просто сидит на месте, пока самец выполняет сложный ритуал ухаживания, она последовательно стимулирует его; эксперименты показали, что каждое действие самца запускается специфическим стимулом. Такая специфика особенно важна в случае близкородственных видов. Как будет показано ниже, их поведенческие особенности, как и морфологические признаки, всегда очень сходны. Прошло слишком мало эволюционного времени для приобретения существенных различий. Однако у таких видов всегда наблюдается какое-либо четкое несовпадение брачных ритуалов, по крайней мере если пространственная (географическое или экологическое) или временная (неодинаковые сезоны размножения) изоляция не устранила в нем необходимости. Например, брачным поведением десятииглая колюшка (Pungitiuspungitius) достаточно похожа на своего трехиглого сородича. Однако у самцов этих видов развилась весьма различная брачная окраска. У десятииглой колюшки самец весной черный как смоль. Этот цвет так же привлекает десятииглых самок, как красный — трехиглых. Вместе с некоторыми незначительными поведенческими различиями неодинаковой окраски достаточно для того, чтобы скрещивание происходило редко.  Рис. 10. Самец десятииглой колюшки показывает самке вход в гнездо. Систематическое исследование проблемы репродуктивной изоляции проводилось на плодовых мушках (Drosophila). Первые результаты показывают, что попытки спаривания между разными видами прерывались на различных стадиях ухаживания, зависящих от используемых в опыте таксонов. Если такое прерывание происходит в серии наблюдений строго в определенный момент, значит, речь идет о специфической реакции, которую не может запустить партнер. Полученные на сегодняшний день результаты показывают, что в некоторых случаях правильный стимул не удается подать самцу, в других же «ошибку» делает самка. Формы ухаживания у различных видов. Формы ухаживания у различных видов очень разнообразны. Ниже приводится несколько примеров. Членистоногие. Первичное обнаружение особи другого пола часто происходит с помощью обоняния. Например, самцы тутового шелкопряда (Bombyxmori) необычайно чувствительны к бомбиколу – половому аттрактанту, выделяемому самкой. Этот аттрактант привлекает самцов на очень больших расстояниях; одной его молекулы достаточно, чтобы вызвать нервный импульс в рецепторной клетке. Сверчки издают разнообразные звуки, которые привлекают самок, а также играют определенную роль в других аспектах поведения при ухаживании. Часто встречаются и зрительные сигналы. Самцы манящего краба (Uca) производят при ухаживании видоспецифические ритуальные движения своей увеличенной клешней. Пауки-скакуны при ухаживании также осуществляют характерные зрительные демонстрации. К наиболее эффектным зрительным демонстрациям относятся брачные сигналы светляков. Летая над полями или лесами, самцы производят вспышки света, носящие видоспецифический характер. Самки реагируют на сигналы самца своего вида короткой вспышкой. Как латентный период, так и сам сигнал характерны для каждого вида, а также зависят от температуры. Самец отвечает на сигнал самки, все больше приближаясь к ней, и пара продолжает обмениваться сигналами до тех пор, пока самец не опускается и не спаривается с самкой. Опытный наблюдатель может привлечь самца, имитируя с помощью миниатюрного фонарика реакцию самки в ответ на вспышки самца. Много работ посвящено ухаживанию у плодовых мушек рода Drosophila. Выявлен ряд элементов поведения (вибрация крыльев, дрожание ножек, сигнализация крылом, кружение и облизывание), которые в различных сочетаниях составляют ритуал ухаживания у разных видов. Рыбы. У разных видов рыб наблюдаются различные формы ухаживания. Наиболее подробно описано поведение одной из тропических аквариумных рыб – Tilapiamelanotheron, у которой выявлены четыре «чисто брачные» формы поведения (которые не встречаются ни при каких других обстоятельствах): быстрые наклоны вниз и вперед, особого рода покачивания головой, покусывание субстрата и неподвижная поза у гнезда. Наиболее выразительная демонстрация в церемонии ухаживания у гуппи – сигмовидная», или S-образная, поза, принимаемая самцом. У меченосцев самец приближается к самке бочком, а также может приближаться к ней, пятясь задом. Амфибии и рептилии. Самцы лягушки-быка занимают определенные территории, откуда слышны их громкие хоры. Самок, очевидно, привлекают такие хоры. Многие виды крокодилов издают громкий рев. Ритуал ухаживания у ящериц Anoliscarolinensis хорошо знаком многим американцам, у которых есть участок при доме. Самец подпрыгивает в определенном ритме, выставляя напоказ ярко-красный горловой мешок (складка кожи, свисающая под подбородком). У большинства видов змей в процедуре ухаживания главную роль играет тактильная стимуляция самки и обонятельная стимуляция самца. В так называемых «брачных танцах», при которых две змеи тесно свиваются одна с другой, как выяснилось, участвуют два самца, и взаимодействие между ними носит, вероятно, агрессивный характер. Птицы. Некоторые из наиболее ярких примеров брачного поведения известны у птиц. Сложные церемонии ухаживания у поганок, чаек, уток, цапель и других птиц служили излюбленными объектами изучения для этологов. Пению птиц и связанным с ним легендам также уделялось значительное внимание. Лоренц изучал различные формы поведения селезней при ухаживании; их можно наблюдать на многих прудах с утками. Селезень наклоняет клюв к воде и выгибает тело кверху; при этом он помахивает клювом и испускает громкий свист, сопровождаемый звуком, напоминающим ворчание. Поза «голова вверх–хвост вниз» сопровождается громким свистом. При демонстрации «вверх-вниз» грудь погружается в воду, а клюв совершает резкий взмах вверх и вперед, поднимая при этом фонтан брызг. Всего у селезней кряквы и других уток, добывающих корм на поверхности воды, известно десять поз, связанных с репродуктивным поведением: 1 – начальное покачивание клюва; 2 – вскидывание головы; 3 – помахивание хвостом; 4 – свист и урчание; 5 – «голова вверх–хвост вверх»; 6 – поворот в сторону самки; 7 – поклоны при плавании; 8 – поворот головы затылком к. самке; 9 – выпячивание груди; 10 – «вверх-вниз». У зеленой кваквы самцы занимают индивидуальные участки. Занявший участок самец прогоняет с него других самцов, прибегая к поведению «полный вперед». Крики самцов привлекают самок. Вначале самок отпугивает поведение «полный вперед», но они не покидают участок, и в конце концов начинается ухаживание в форме «щелканья» и «вытягивания шеи». Образовав пару, самец и самка летают по территории, иногда демонстрируя интенсивный машущий полет. Последующее поведение ведет ко все большему сближению, пока не произойдет копуляция. У многих птиц пение служит для того, чтобы прогонять других самцов с занятого участка и привлекать самок. Характер песни у некоторых видов столь же специфичен, как и их внешние признаки. Млекопитающие. Важную роль в регуляции брачного поведения млекопитающих поведения у многих видов играет обоняние. Широко распространены такие действия, как обследование аногенитальной области и обнюхивание мочевых меток. У многих видов самцы, почуяв запах самки, проявляют реакцию Флемена, выражающуюся в вытягивании шеи и приподнимании верхней губы. Эта реакция, по-видимому, скорее облегчает восприятие запаха, чем служит демонстрацией. Самки млекопитающих нередко побуждают самца к садке, иногда приближаясь к нему, обнюхивая его и облизывая, а часто убегая от него. Во многих случаях бегство самок от самца, по-видимому, служит для того, чтобы привлечь его, а не для того, чтобы действительно убежать. У дельфинов-афалин ритуал ухаживания включает вокализации, покусывание партнера, обнюхивание его половых органов, трения друг о друга, поглаживания плавниками, выставления напоказ белой нижней поверхности тела, прыжков, погони друг за другом и «бодания». ЗаключениеЭтот очень краткий и фрагментарный обзор, возможно, достаточен для того, чтобы продемонстрировать сложную природу поведенческих схем, обеспечивающих сотрудничество партнеров по спариванию. Было показано, что следует различать типы функций, выполняемых ухаживанием. Это не означает, что каждое отдельное действие при ухаживании служит только одной из этих целей. Например, зигзагообразный танец самца колюшки, безусловно, обеспечивает синхронизацию, соблазнение, ориентацию и репродуктивную изоляцию, хотя различия в брачной окраске трехиглой и десятииглой колюшек объяснимы только с точки зрения изоляции. Известны также действия, связанные с синхронизацией и соблазнением, но не с ориентацией: например, самки бабочки сатира могут быть «настроены» и соблазнены ухаживанием одного самца, а затем спариться с другим. Это означает, что первый самец не ориентировал реакцию самки в свою сторону. Аналогичным образом у голубей настойчивое воркование самца не столько ориентирует на него голубку, сколько способствует овуляции ее гонад. У различных видов близкородственных дарвиновых вьюрков с островов Галапагос, как выяснилось, почти идентичные способы ухаживания за самками, однако межвидового скрещивания не происходит. Здесь репродуктивная изоляция обеспечивается отчасти различными экологическими нишами, а отчасти тем, что каждый партнер специфически реагирует на свойственную только его виду форму клюва, которая связана с особенностями потребляемой пищи. В данном случае ухаживание не имеет ничего общего с репродуктивной изоляцией и служит совсем другим задачам. Брачное поведение завершается актом совокупления (копуляции), обеспечивающего оплодотворение яиц. К основным типам оплодотворения относятся внутреннее и наружное. Формы копуляции почти столь же разнообразны, как и формы ухаживания. Список литературыГодфруа «Что такое психология» том 1, Москва, изд. «Мир», 1999г.; Дьюсбери Д. 1981. Поведение животных: Сравнительные аспекты. – М.: Мир, 1981.; К. Лоренц «Агрессия (так называемое «зло»)», Москва, изд. группа «ПРОГРЕСС», «УНИВЕРС» 1994г.; Пианка Э. Эволюционная экология. – М.: Мир, 1981.; Н. Тинберген «Социальное поведение животных»; К. Фабри «Основы зоопсихологии», Москва, Умк «Психология», 2001г.  |