генетика микроорганизмов. Генетика микроорганизмов. Фенотип и генотип микроорганизмов
 Скачать 221.3 Kb. Скачать 221.3 Kb.
|

. Оба штамма в течение ночи выращивали вместе на полноценной среде. Затем смешанную культуру центрифугировали, отмывали от полноценной среды и высевали на минимальную питательную среду. На минимальной питательной среде без метионина, биотина, треонина и лейцина появились прототрофные колонии Met+ Bio+ Thr+ Leu+ с частотой около 1 на каждые 107 клеток.
и не способный синтезировать треонин и лейцин штамм Thr«Генетика микроорганизмов.» Фенотип и генотип микроорганизмов. Материальной основой наследственности бактерий является ДНК. По сравнению с геном эукариотов геном бактерий устроен более просто - это молекула ДНК, замкнутая в кольцо, которое прикреплено к одной из мезосом. Кроме хромосомы, у бактерий имеются внехромосомные генетические элементы - плазмиды. Это молекулы ДНК, которые или находятся вне хромосомы, в автономном состоянии, в виде колец, прикрепленных к мезосомам, или встроены в хромосому (интегрированное состояние). Плазмиды придают бактерии дополнительные наследственные признаки, но не являются обязательными для нее. Генотип — это общая сумма генов микроба. В отношении микроорганизмов "генотип" означает то же, что "геном". Это возможные способности клетки. Фенотип — это весь комплекс свойств микроба, проявление генотипа в определенных, конкретных условиях существования. Видимое проявление способностей клетки. Строение и размножение бактерий. Химический состав клеток бактерий в основном такой же, как и клеток высокоорганизованных организмов. Клетки бактерий окружены оболочкой, внутри которой находятся цитоплазма, ядерный аппарат, рибосомы, ферменты и другие включения. В отличие от клеток эукариот у них отсутствуют митохондрии, пластинчатый комплекс и эндоплазматическая сеть. В центральной части цитоплазмы бактерий расположены ядерный аппарат — нуклеоид и плазмиды. Ядро прокариот называется нуклеотидом потому, что оно в отличие от эукариот не изолировано от цитоплазмы мембраной и представлено одной очень длинной молекулой ДНК. Кроме нуклеотида в цитоплазме большинства бактерий содержатся так называемые внехромосомные факторы, получившие название плазмид. Плазмиды представляют собой кольцевые молекулы ДНК, обладают свойствами репликона — могут реплицироваться с помощью ферментов клетки бактерии независимо от основной хромосомы. Плазмида включает последовательность из одного или нескольких генов. Плазмиды реплицируются в цитоплазме автономно и передаются при делении дочерним клеткам. При размножении клетки бактерии наиболее ответственным является процесс воспроизведения нуклеотида. В нуклеотиде ДНК суперспирализована и плотно уложена. Один конец ДНК прикреплен к клеточной мембране. Связь с клеточной мембраной необходима как для процесса репликации ДНК, так и для четкого разделения вновь образовавшихся дочерних молекул ДНК. Репликация ДНК у микроорганизмов происходит так же, как и у высших организмов. В репликации участвуют ферменты ДНК- полимеразы. К моменту завершения цикла репликации ДНК точки прикрепления дочерних ДНК отодвигаются благодаря активному росту участка бактериальной мембраны между ними. В результате сложного комплекса процессов образуется межклеточная перегородка. В период репликации ДНК и образования перегородки клетка непрерывно растет, идет формирование рибосом и других соединений. На определенной стадии дочерние клетки отделяются друг от друга. Каждая дочерняя клетка имеет такой же набор генетической информации, какой был в исходной бактериальной клетке. Строение и размножение вирусов. Вирус (от латинского virus – яд) — это особая форма неклеточного организма, который обладает геномом РНК или ДНК, но не имеет своего синтезирующего аппарата. Вирусы способны производить себе подобных лишь в клетках других, более развитых существ. История открытия. В 1852 году русский ботаник Д. И. Ивановский впервые получил инфекционный экстракт из растений табака, пораженных мозаичной болезнью. Когда такой экстракт пропустили через фильтр, способный задерживать бактерии, отфильтрованная жидкость все еще сохраняла инфекционные свойства. Сами частицы все еще оставались неуловимыми и загадочными, потому что они были слишком малы, чтобы их можно было увидеть с помощью светового микроскопа. Поэтому-то вирусы и оказались в числе первых биологических структур, которые были исследованы в электронном микроскопе сразу же после его изобретения. Строение вирусов. Учитывая тот факт, что в состав организма вируса входит либо РНК, либо ДНК, эти вирусы разделяют на два типа: РНК-содержащие и ДНК-содержащие. В свою очередь каждый из этих типов подразделяется на одноцепочечные и двухцепочечные. Как правило, РНК-содержащие вирусы имеют линейную структуру, а ДНК-содержащие - кольцевую и линейную структуры. Особенность вирусов — они могут себя проявлять, размножаться и мутировать только находясь в клетках чужих организмов. Вне клеток они никак себя не проявляют. Во внешней среде форма многих из вирусов напоминает кристалл, а размеры их в диаметре варьируются от 20 до 300 нм.  Капсид — это белковая структура, в которую заключен генетический материал вируса. Вирион — полностью сформированная инфекционная частица. 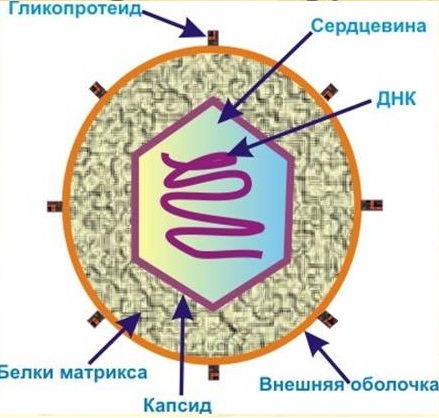 Простые вирусы состоят из белков и нуклеиновой кислоты. Наиболее важная часть вирусной частицы – нуклеиновая кислота – носитель генетической информации. У разных вирусов обнаружен, лишь один тип – или ДНК, или РНК, что положено в основу их классификации. Второй обязательный компонент вириона – белки, которые отличаются у разных вирусов, что позволяет распознавать их с помощью иммунологических реакций. Более сложные по структуре вирусы, кроме белков и нуклеиновых кислот, содержат углеводы, липиды. Для каждой группы вирусов характерен свой набор белков, жиров, углеводов и нуклеиновых кислот. Некоторые вирусы содержат в своем составе ферменты. У сложно устроенных вирусов сердцевина в виде туго свернутой спирали покрыта одной или несколькими внешними оболочками, в состав которых входят различные вещества. Размножение вирусов:
Встреча вируса с клетками начинается с его адсорбции, т.е. прикрепления к клеточной стенке, плазматической мембране клетки. Причем каждый вирион способен прикрепляться лишь к определенным клеткам, имеющим специальные рецепторы. На одной клетке могут адсорбироваться десятки и даже сотни вирионов. Затем клетка как бы «втягивает» прикрепившиеся вирионы внутрь. Следующий этап – «раздевание» проникших внутрь клеток вирионов. Для этой цели используется имеющийся в клетках комплекс специальных ферментов, которые растворяют белковый чехол вируса и освобождают его нуклеиновую кислоту. Последняя по клеточным каналам проникает в ядро клетки или остается в цитоплазме клетки. Она не только руководит размножением вируса, но и определяет его наследственные свойства. Нуклеиновая кислота вируса подавляет собственный обмен клетки и направляет его на производство новых компонентов вируса. С помощью полимераз снимаются копии родительской нуклеиновой кислоты . Часть вновь образовавшихся копий соединяется с рибосомами, на которых осуществляется синтез вирусных белков. После того как в зараженной клетке накопится достаточное количество компонентов вируса, начинается процесс композиции, т.е сборка вирионов. Процесс этот происходит обычно вблизи клеточных оболочек, иногда принимающих в нем непосредственное участие. Последним этапом взаимодействия вирусов с клетками является выход новых дочерних вирионов из клетки. До гибели клеток эти вирусы успевают проделать несколько циклов размножения, постепенно истощая синтетические ресурсы клеток. В отдельных случаях вирусы могут накапливаться внутри клеток, образуя кристаллоподобные скопления — тельца включений. Способы передачи наследственного материала у бактерий и вирусов. Трансформация. Трансформация заключается в том, что ДНК, выделенная из бактерий в свободной растворимой форме, передается бактерии-реципиенту. При трансформации рекомбинация происходит, если ДНК бактерий родственны друг другу. В этом случае возможен обмен гомологичных участков собственной и проникшей извне ДНК. Впервые явление трансформации описал Фредерик Гриффите (1928). Он вводил мышам живой невирулентный бескапсульный R-штамм пневмококка и одновременно убитый вирулентный капсульный S-штамм пневмококка. Из крови погибших мышей был выделен вирулентный пневмококк, имеющий капсулу убитого S-штамма пневмококка. Таким образом, убитый S-штамм пневмококка передал наследственную способность капсулообразования R-штамму пневмококка. Освальд Эвери, Колин Маклеод и Маклин Маккарти (1944) доказали, что трансформирующим агентом в этом случае является ДНК. Путем трансформации могут быть перенесены различные признаки: капсулообразование, устойчивость к антибиотикам, синтез ферментов. Конъюгация. Конъюгация бактерий состоит в переходе генетического материала (ДНК) из клетки-донора («мужской») в клетку-реципиент («женскую») при контакте клеток между собой. Открыт в 1946 году Джошуа Ледербергом и Эдуардом Тайтемом. Они провели следующий эксперимент. Были отобраны два ауксотрофных мутантных штамма Е. coli К-12: не способный синтезировать метионин и биотин штамм Met |