ginekologia_otvety_na_ekz (копия). Гинекология современные представления о нейрогуморальной регуляции менструального цикла. Показатели норального менструального цикла
 Скачать 239.42 Kb. Скачать 239.42 Kb.
|

|
Гинекология 1. СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О НЕЙРОГУМОРАЛЬНОЙ РЕГУЛЯЦИИ МЕНСТРУАЛЬНОГО ЦИКЛА. ПОКАЗАТЕЛИ НОРАЛЬНОГО МЕНСТРУАЛЬНОГО ЦИКЛА. Основной функцией репродуктивной системы (РС) является воспроизводство, т.е. продолжение биологического вида. Оптимальной функциональной активности РС достигает к 16 - 18 годам – возрасту, когда организм находится в оптимальных условиях для зачатия, вынашивания беременности и вскармливания ребенка. Особенностью РС является ее постепенное угасание. С 45 лет угасает генеративная функция, с 50 лет – менструальная и затем – гормональная. Готовность женского организма к наступлению беременности внешне проявляется в виде менструальных кровотечений, указывающих на циклические сдвиги, происходящие в организме в целом и, особенно, в половых органах. Эти периодические изменения объединены под названием менструального цикла. Каждый нормальный менструальный цикл является подготовкой организма женщины к беременности.  ВНЕШНЯЯ СРЕДА Схема 1. Функциональная структура регуляции репродуктивной системы.                            Андрогены Эстрогены Прогестерон Ингибин ЯИЧНИКИ ФР ПрЛ ФСГ ЛГ ГР ПЕРЕДНЯЯ ДОЛЯ ГИПОФИЗА ГОНАДОТРОПИН-РИЛИЗИНГ-ГОРМОН КОРА ГОЛОВНОГО МОЗГА ЭКСТРАГИПОТАЛАМИЧЕСКИЕ СТРУКТУРЫ    ОРГАНЫ И ТКАНИ-МИШЕНИ Регуляция менструальной функции осуществляется сложным нейро–гуморальным путем. Циклические процессы происходят при обязательном участии пяти важнейших звеньев (уровней) регуляции (схема 1): кора головного мозга и экстрагипоталамические церебральные структуры (мозолистое тело, таламус, шишковидное тело, латеральное и медиальное коленчатые тела и др.); подкорковые центры, расположенные преимущественно в области гипоталамуса; придаток мозга – гипофиз; половые железы – яичники; периферические органы и ткани-мишени (маточные трубы, матка, влагалище, молочные железы, кожа, волосяные фолликулы, потовые, сальные железы, жировая ткань, кости, мышцы). Как и все эндокринные системы в организме, репродуктивная функционирует по механизму отрицательной обратной связи, когда при снижении уровня периферических гормонов усиливается синтез и выделение тропных гормонов гипофиза. Однако, для РС характерна еще и положительная обратная связь, когда в ответ на максимальное повышение уровня эстрадиола в преовуляторном периоде увеличивается синтез и выделение гонадотропинов. Ведущее место в регуляции РС принадлежит гипоталамо-гипофизарной системе, которая определяет состояние и функционирование всей эндокринной системы. В свою очередь, кора головного мозга и экстрагипоталамические структуры, получающие всю информация из внешней и внутренней среды, осуществляют контроль над гипоталамо-гипофизарной системой посредством нейромедиаторов, т.е. передатчиков нервного импульса на нейросекреторные ядра гипоталамуса. Гипоталамус – высший вегетативный центр, координирующий функции всех внутренних систем, поддерживающих внутренний гомеостаз в организме. Под контролем гипоталамуса находится гипофиз и регуляция эндокринных желез: гонад, щитовидной железы, надпочечников. Гипоталамус представляет собой скопление нервных клеток, обладающих нейросекреторной активностью. Местом синтеза гипофизотропных рилизинг-гормонов (РГ) или либеринов являются вентро- и дорсомедиальные, аркуатные ядра гипоталамуса. Основным гипоталамическим РГ, регулирующим репродуктивную функцию на уровне гонадотрофов гипофиза, является гонадотропин-рилизинг-гормон (Гн-РГ), представляющий собой декапептид. Хотя первоначально считалось, что для лютеинизирующего (ЛГ) и фолликулостимулирующего (ФСГ) гормонов гипофиза существуют отдельные РГ, современный взгляд состоит в том, что Гн-РГ является единственным универсальным гонадотропин-рилизинг-гормоном. Вариации ответа гипофиза в виде синтеза ЛГ или ФСГ считаются вызванными эффектами обратной связи стероидных гормонов яичника на уровне гипоталамо-гипофизарной оси. Секреция Гн-РГ в гипоталамусе подвергается влиянию нейронов, расположенных в различных регионах головного мозга, чьи терминалы заканчиваются в аркуатном ядре. Такие нейротрансмиттеры, как эпинефрин и норэпинефрин, повышают высвобождение Гн-РГ, в то время, как дофамин, серотонин и эндогенные опиоидные пептиды, напротив, являются ингибиторами секреции Гн-РГ. Главный эффект Гн-РГ заключается в его действии на гонадотрофные клетки передней доли гипофиза, что приводит к стимуляции процессов секреции молекул ЛГ и ФСГ. Секреция Гн-РГ генетически запрограммирована и происходит в определенном пульсирующем режиме, примерно 1 раз в час (цирхоральный режим). Через портальную кровеносную систему, объединяющую гипоталамус и гипофиз, Гн-РГ поступает в гипофиз. Особенностью этой кровеносной системы является возможность тока крови в ней в обе стороны, что важно для осуществления механизмов обратной связи (ультракороткой и короткой). Гипофиз – место синтеза и выделения всех тропных гормонов, непосредственно регулирующих функцию периферических эндокринных желёз. Местом синтеза гонадотропных гормонов (ЛГ и ФСГ), пролактина (ПРЛ) является передняя доля гипофиза – аденогипофиз. ЛГ и ФСГ подобно тиротропину и хорионическому гонадотропину (ХГ) являются гликопротеинами, состоящими из двух полипептидных цепей, обозначаемых α и β. α-цепи всех четырех гормонов идентичны, β-цепи – уникальны и обеспечивают специфичную биологическую активность для каждого гормона. ЛГ и ФСГ определяют первые этапы синтеза стероидов в яичниках путем взаимодействия со специфическими рецепторами клеточной мембраны органа-мишени – яичников. Эффективность гормональной регуляции определяется как количеством активного гормона, так и уровнем содержания рецепторов в клетке-мишени. Под влиянием ФСГ происходит рост и созревание фолликула. В этот процесс входит: пролиферация клеток гранулёзы; синтез рецепторов ЛГ на поверхности клеток гранулёзы; синтез ферментных систем – ароматаз, участвующих, в метаболизме андрогенов в эстрогены; содействие овуляции (совместно с ЛГ). Под влиянием ЛГ происходят: первые этапы синтеза андрогенов в клетках теки фолликула; синтез эстрадиола в клетках гранулёзы доминантного фолликула; стимуляция овуляции; синтез прогестерона в лютеинизированных клетках гранулёзы (жёлтом теле). Пролактин синтезируется клетками аденогипофиза (лактотрофами), является одноцепочечным белком. Рецепторы пролактина широко распространены во всем организме. Фактически трудно найти ткань, которая не экспрессирует молекулу рецептора. В настоящее время идентифицировано более 300 эффектов, включая эффекты на водный и солевой баланс, рост и развитие, эндокринологию и метаболизм, мозг и поведение, репродукцию и иммунную систему. Гипоталамический контроль в регуляции синтеза ПРЛ проявляется тормозящим эффектом дофамина. Рилизинг-гормон к ПРЛ до настоящего времени не выделен, но известно, что тиролиберин стимулирует синтез ПРЛ лактотрофами гипофиза. Яичники – парные женские половые железы, выполняющие генеративную и эндокринную функции. Они состоят из коркового и мозгового вещества. В корковом слое располагаются фолликулы различной степени зрелости. Яичники секретируют разнообразные по химической структуре и физиологической активности гормоны. Условно все гормоны яичников можно разделить на стероидные и белково-пептидные. К первым относят эстрон, эстрадиол, прогестерон и андрогены, такие как дегидроэпиандростендион, андростендион, тестостерон и дигидротестостерон. Белково-пептидные гормоны представлены фолликулостатином, активином, ингибином, фолликулярно-регулирующим протеином, ингибитором созревания ооцитов, релаксином и др. Помимо этого яичники секретируют такие гормоноиды, как аденозин, катехоламины, эйкозаноиды, γ-аминомасляную кислоту, а также факторы роста. Стероидпродуцирующими структурами яичников являются клетки гранулезы, теки и в меньшей степени – строма. Клетки теки являются главным источником андрогенов и незначительного количества эстрогенов. Основное место синтеза эстрадиола – клетки гранулёзы. Прогестерон синтезируется в клетках теки и лютеинизированной гранулезы. Субстратом для всех стероидов служит холестерин. Первые этапы синтеза стероидов детерминированы гонадотропинами. Под влияние ЛГ в клетках теки начинается синтез андрогенов, основная часть которых поступает в клетки гранулезы, где под влиянием ароматаз (их синтез индуцируется ФСГ) происходит превращение андрогенов в эстрогены. Схема 2. Синтез эстрогенов в яичниках (двуклеточная теория). КРОВЬ  ФОЛЛИКУЛЯРНАЯ ЖИДКОСТЬ ФОЛЛИКУЛЯРНАЯ ЖИДКОСТЬ  Холестерин Холестерин ЛГ ФСГ ЛГ ФСГ    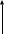 //////////////////////////////////////////////////////////// Холестерол Прегненолон Андростендион Эстрадиол Эстрон ароматаза Андростендион Клетка теки Клетка гранулезы Базальная мембрана Факторы роста (ФР) – биологически активные вещества, стимулирующие или ингибирующие дифференцировку клеток – являются основными переносчиками митогенного сигнала деления клеток. ФР обладают аутокринным, паракринным, интракринным и эндокринными эффектами. Важную роль в физиологии РС играют следующие ФР: Инсулиноподобные факторы роста I и II (ИПРФ-I и ИПРФ-II) вырабатываются под влиянием гормона роста. Местом синтеза в яичниках являются клетки гранулёзы. Они потенцируют действие гонадотропинов, усиливая их стероидогенную активность. Эпидермальный фактор роста (ЭФР) относится к наиболее мощным стимуляторам клеточной пролиферации. Он обнаружен в клетках гранулёзы, стромальных клетках эндометрия, молочных железах и других тканях, обладает онкогенным эффектом в эстрогензависимых тканях (эндометрий, молочные железы). Трансформирующие факторы роста (α-ТФР и β-ТФР) также обладают сильным митогенным эффектом, участвуют в росте и созревании фолликулов, пролиферации клеток гранулезы. ТФР обладает онкогенным действием. Не исключена роль андрогенов и инсулина в индукции синтеза α-ТФР. Сосудистый эндотелиальный фактор роста (СЭФР) обладает мощным митогенным эффектом для эндотелиальных клеток, усиливает проницаемость сосудов, участвует в ангиогенезе. Показана роль СЭФР в функции клеток гранулезы преовуляторного фолликула. Ингибин – белковое вещество, образующееся в клетках гранулезы фолликула. Ингибин участвует в регуляции секреции ФСГ, тормозя ее, подобно эстрадиолу, по сходному механизму обратных связей. Образование ингибина возрастает к овуляции под влиянием ФСГ, а достигнув максимума, тормозит его выделение. Процесс фолликулогенеза происходит в яичниках непрерывно с антенатального периода до постменопаузы. Основная масса фолликулов (около 90%) подвергается атрезии, а остальные проходят полный цикл развития – от примордиального до преовуляторного, овулируют и превращаются в желтое тело. В процессе атрезии фолликулов важная роль отводится апоптозу (программируемой гибели клеток) – биологическому процессу, в результате которого полностью рассасываются клетки под влиянием собственного лизосомального аппарата. В яичнике женщины репродуктивного возраста фолликулы находятся на различных стадиях зрелости. Примордиальные фолликулы характеризуются одним слоем плоских преграгулёзных клеток, небольшим ооцитом, клетки теки отсутствуют. Первичные преантральные фолликулы характеризуются одним слоем клеток гранулезы, на этой стадии начинается образование клеток теки и увеличение ооцита. Вторичные преантральные фолликулы характеризуются 2-8 слоями клеток гранулезы и полностью сформированным слоем клеток теки. Антральные фолликулы имеют в центре полость, заполненную жидкостью; диаметр фолликулов к началу менструального цикла составляет около 3 мм, они имеют тенденцию к быстрому росту в ранней фолликулярной фазе. Преовуляторный фолликул достигает до 18 – 25 мм в диаметре, имеет много слоёв клеток гранулёзы, ооцит располагается на одной из сторон полости фолликула. После овуляции на месте фолликула образуется желтое тело. Рост фолликула от покоящегося примордиального до преовуляторного носит последовательный и непрерывный характер и занимает около 85 суток. Рост до стадии малых антральных фолликулов является гормонально независимым и регулируется местными яичниковыми факторами. Под влиянием каких факторов происходит выход фолликулов из покоящегося примордиального состояния, до настоящего времени не выяснено. Число растущих фолликулов зависит от возраста женщины. Так, в 24 – 25 лет число растущих фолликулов, покидающих примордиальный пул, составляет около 37 в день, в 34 – 35 лет – 11, а к 44 – 45 годам – только 2. Гонадотропинзависимый рост овариальных фолликулов начинается с повышения уровня ФСГ в конце предыдущего менструального цикла. Увеличение синтеза и выделения ФСГ гипофизом происходит по механизму отрицательной обратной связи в ответ на снижение уровня эстрадиола, прогестерона и ингибина при регрессе желтого тела. При этом формируется пул растущих антральных фолликулов, из которых выбирается доминантный – преовуляторный фолликул в последующем цикле. Стадии фолликулогенеза от конца лютеиновой фазы предыдущего цикла делят на следующие этапы: Набор когорты антральных фолликулов, Селекция доминантного фолликула, Овуляция с разрывом фолликула и выходом ооцита, Формирование и функционирование желтого тела. Повышение уровня ФСГ в конце лютеиновой фазы предыдущего цикла стимулирует рост антральных фолликулов с 1-3 до 5-6 мм в ранней фолликулярной фазе цикла. С ростом фолликула резко уменьшается синтез ЭФР клетками гранулезы, который блокирует выработку ингибина и чувствительность клеток гранулезы к ФСГ. Особое значение имеет базальный уровень ФСГ на 2 – 4 день менструального цикла. Этот показатель отражает минимальный уровень ФСГ, необходимый для формирования пула антральных фолликулов, и способность клеток гранулезы синтезировать ингибин и эстрадиол в ответ на действие ФСГ. Увеличение базального уровня ФСГ отражает уменьшение пула антральных фолликулов и, соответственно, уменьшение выработки ими ингибина В и эстрадиола. Согласно двуклеточной теории стероидогенеза в яичниках, ЛГ стимулирует синтез из холестерина андрогенов в клетках теки, тогда как ФСГ стимулирует ароматизацию андрогенов в эстрогены в клетках гранулезы (схема 2). В ранней фолликулярной фазе до 6 – 7 дня менструального цикла рост фолликулов зависит от ФСГ, их размер составляет в диаметре от 5 до 10 мм. Число растущих фолликулов четко коррелирует с возрастом женщины. В этот период ФСГ стимулирует пролиферацию и дифференцировку клеток гранулёзы, синтез в них ЛГ-рецепторов, активацию ароматаз и продукцию ингибина В. ЛГ в ранней фолликулярной фазе влияет преимущественно на синтез андрогенов в клетках теки и имеет мало рецепторов на клетках гранулезы. Максимального значения уровень ФСГ достигает к 5 – 6 дню цикла, после чего он снижается и затем повышается одновременно с ЛГ. Считается, что селекция доминантного фолликула происходит в период роста пула антральных фолликулов на 5 – 7 день менструального цикла при размере фолликулов 5 – 10 мм. Доминантным становится фолликул наибольшего диаметра, с наибольшим числом клеток гранулезы и рецепторов ФСГ, что позволяет синтезировать наибольшее количество ингибина и эстрадиола. Доминантный фолликул может сохранить способность к дальнейшему росту и синтезу эстрадиола к моменту снижения уровня ФСГ. Увеличение синтеза эстрадиола по механизму положительной обратной связи все больше повышает выработку ЛГ, что в свою очередь увеличивает синтез эстрадиола в клетках гранулёзы. В быстром росте лидирующего фолликула играют роль возрастающие концентрации эстрадиола и ИПФР, синтез которых в клетках гранулезы активируется под влиянием гормона роста и гонадотропинов. Синтез ингибина клетками гранулезы преовуляторного фолликула приводит к снижению выработки ФСГ гипофизом. В остальных фолликулах, в которых синтез эстрадиола остается ФСГ-зависимым, снижающиеся дозы ФСГ вызывают процесс атрезии, механизм которого связан с апоптозом. Состав фолликулярной жидкости доминантного и атретичных фолликулов различный. Доминантный фолликул характеризуется высокой концентрацией эстрадиола и высоким значением отношения уровня эстрадиола к тестостерону, что свидетельствует о высокой активности ароматаз, высокой концентрации ИПФР. В атретичном фолликуле, наоборот, высока концентрация тестостерона, отношение эстрадиола к тестостерону низкое, снижена концентрация ИПФР. Процесс овуляции осуществляется при достижении максимального уровня эстрадиола в преовуляторном фолликуле, который по механизму положительной обратной связи стимулирует овуляторный выброс ЛГ и ФСГ гипофизом. Овуляция происходит через 10 – 12 часов после пика ЛГ или через 24 – 36 часов после пика эстрадиола. Базальная мембрана фолликула разрывается под влиянием различных ферментов и биологически активных субстанций в клетках гранулезы: протеолитических ферментов, плазмина, гистамина, коллагеназы, простагландинов, окситоцина и релаксина, под влиянием которых истончается стенка фолликула. После овуляции клетки гранулезы подвергаются лютеинизации с образованием желтого тела – временного эндокринного органа, секретирующего прогестерон под влиянием ЛГ в течение 14 дней. В отсутствие беременности желтое тело подвергается лютеолизу и превращается в фиброзный рубец, белое тело. К органам-мишеням действия половых гормонов относятся матка, влагалище, молочные железы, волосяные фолликулы, кожа, жировая ткань, мышцы, кости. Клетки названных тканей и органов содержат рецепторы к половым гормонам. Изменения гормонального фона (содержание в крови эстрогенов и прогестерона в разные дни овариального цикла) оказывают прямое влияние на состояние эндометрия, слизистой оболочки маточных труб, цервикального канала и влагалища. В эндометрии различают функциональный (отпадающий при менструации) и базальный (сохраняющийся при менструации) слои. Базальный слой слизистой оболочки матки прилежит к мышечному и состоит из желез и стромы, его толщина составляет 1 – 1,5 мм. В этом слое расположены самые нижние отделы и дно желез эндометрия. С базального слоя начинается процесс эпителизации и регенерации функционального слоя после менструаций, абортов, родов. Циклические изменения в строении и функции матки в целом и, особенно, в строении и функции эндометрия под воздействием яичниковых гормонов, описываются под названием маточного цикла. В течение маточного цикла наблюдается последовательная смена четырех фаз циклических изменений в эндометрии: Пролиферации; Секреции; Десквамации; Регенерации. Фаза пролиферации эндометрия начинается после завершения регенерации слизистой оболочки, отторгнувшейся во время предшествующей менструации и длится до момента овуляции. Пролиферативная фаза заключается в росте эндометрия под влиянием эстрогенов, фиксируются множественные митозы. Эндометриальные железы сначала узкие и трубчатые, постепенно увеличиваются и начинают слегка извиваться. Происходит врастание спиральных артерий из базального слоя. Фаза секреции продолжается от овуляции до начала менструации. В ранней стадии фазы секреции эпителиальные клетки под воздействием прогестерона прекращают деление и гипертрофируются. Маточные железы расширяются, становятся более разветвленными. Железистые клетки начинают секретировать гликоген, гликопротеины, липиды, муцин. В последующем просветы желез расширяются, стенки их становятся складчатыми. Спиральные артерии резко извилисты, образуют клубки. На 21-й 22-й дни (при 28-дневном цикле) эндометриальная строма становится отечной, возникает децидуальная реакция. Именно к этому времени эндометрий становится подготовленным к имплантации. В случае отсутствии оплодотворения и имплантации происходит редукция желтого тела, что приводит к снижению эстрогенов и прогестерона в плазме. Уменьшение содержания стероидных гормонов яичников приводит к скручиванию, склерозированию и уменьшению просвета спиральных артерий. К концу поздней стадии фазы секреции вены в поверхностных слоях эндометрия расширены, переполнены кровью, в них образуются тромбы. В результате происходит ухудшение кровотока в функциональном слое эндометрия – ишемия, что приводит к гибели клеток и к отторжению функционального слоя и кровотечению (фаза десквамации). Менструальная жидкость состоит из крови и поверхностных эндометриальных тканей. После возобновления секреции эстрадиола фолликулами яичников из тканей базального слоя происходит регенерация. Железы слизистой оболочки канала шейки матки, секретирующие слизь, также подвергаются циклическим изменениям. После менструации количество слизи ограничено и она имеет вязкую консистенцию. В ответ на растущие уровни эстрадиола, секретируемого фолликулом во время более поздней фолликулярной фазы, количество слизи возрастает примерно в 30 раз. Слизь также меняет качество и становится более водянистой и пластичной. Прогестерон реверсирует эффекты эстрогенов на уровне слизистой шейки матки. Вагинальные эпителиальные клетки также подвергаются влиянию эстрогенов и прогестерона. Во время ранней фолликулярной фазы слущенные влагалищные эпителиальные клетки являются базофильными и имеют везикулярные ядра. В ответ на повышающиеся уровни эстрадиола во второй половине фолликулярной фазы начинают доминировать ацидофильные клетки с пикнотическими ядрами. Во время лютеиновой фазы, когда повышается уровень прогестерона, доля ацидофильных клеток снижается. |