02 Гистология. Гистология животных Эпителиальная ткань животных 3
 Скачать 2.81 Mb. Скачать 2.81 Mb.
|

|
Поперечнополосатая мышечная ткань В 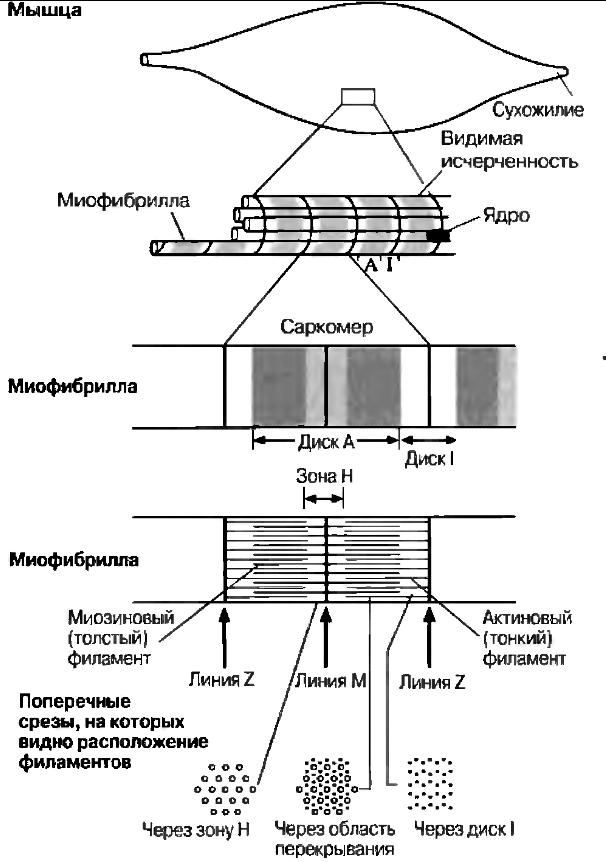 нешний вид продольного среза мышцы (или выделенных из нее отдельных мышечных волокон) объясняет, почему ее называют поперечнополосатой. Однако причины такой характерной исчерченности становятся понятными только при электронномикроскопическом анализе. Поперечнополосатая мышца состоит из множества вытянутых клеток, называемых из-за своей формы мышечными волокнами. Длина их достигает нескольких сантиметров при диаметре 0,01—0,1 мм (диаметр обычной клетки около 0,02 мм). Волокна имеют цилиндрическую форму и расположены параллельно друг другу. Каждое волокно многоядерное — этим оно отличается от мышечных клеток другого типа. Ядра в волокне расположены около его поверхности. Пучки мышечных волокон окружены коллагеновыми волокнами и соединительной тканью; между волокнами тоже находится коллаген. Каждое волокно окружено мембраной — сарколеммой, которая по своему строению очень напоминает обычную плазматическую мембрану. нешний вид продольного среза мышцы (или выделенных из нее отдельных мышечных волокон) объясняет, почему ее называют поперечнополосатой. Однако причины такой характерной исчерченности становятся понятными только при электронномикроскопическом анализе. Поперечнополосатая мышца состоит из множества вытянутых клеток, называемых из-за своей формы мышечными волокнами. Длина их достигает нескольких сантиметров при диаметре 0,01—0,1 мм (диаметр обычной клетки около 0,02 мм). Волокна имеют цилиндрическую форму и расположены параллельно друг другу. Каждое волокно многоядерное — этим оно отличается от мышечных клеток другого типа. Ядра в волокне расположены около его поверхности. Пучки мышечных волокон окружены коллагеновыми волокнами и соединительной тканью; между волокнами тоже находится коллаген. Каждое волокно окружено мембраной — сарколеммой, которая по своему строению очень напоминает обычную плазматическую мембрану.В световом микроскопе внутри мышечного волокна можно различить множество тонких миофибрилл (от греч. myos-- мышца и fibrilla — волоконце), которые и создают характерную поперечную исчерченность. Ультраструктура поперечнополосатых мышц Понять механизм сокращения мышцы помогает исследование ее ультраструктуры в электронном микроскопе, который позполяет детально рассмотреть миофибриллы: их диаметр близок к 1 мкм (у волокна в 100 раз больше). Их пересекают темные полосы, которые и определяют исчерченность мышечного волокна. Более детальное изучение показывает, что миофибрилла образована параллельно проходящими нитями двух типов — тонкими и толстыми миофиламентами. Они состоят из разных белков: первые — из актина, вторые — из миозина. Местами тонкие миофиламенты заходят в промежутки между толстыми, как частично переплетенные пальцы двух ладоней. Эти участки перекрывания и соответствуют темным полосам, что тоже легко продемонстрировать на пальцах. Темные полосы принято называть зонами, или дисками А. а светлые между ними — зонами, или дисками I. В последних присутствуют только актиновые миофиламенты. В зоне А можно различить среднюю зону, где нет перекрывания, т. е. проходят лишь миозиновые нити. Ее называют зоной Н. Наконец, зона Н разделяется пополам линией М. а зона I —линией Z. Как выяснилось, функциональной единицей мышцы является способный сокращаться отрезок миофибриллы между линиями Z, названный саркомером. Миофибрилла, а, cледовательно и мышечное волокно в целом, состоит из тысяч саркомеров. Механизм мышечного сокращения; теория скользящих нитей Когда была выявлена ультраструктура миофибрилл, две независимые группы исследователей (X. Хаксли/Дж. Хансон и А. Хаксли/Р. Нидергерке) предложили гипотезу мышечного сокращения, основанную на скольжении относитель но друг друга актиновых и миозиновых нитей. Ее легко понять, вдвигая пальцы одной руки между пальцами другой: если считать обе ладони эквивалентом одного саркомера, (к оглавлению) то эта система будет укорачиваться, т. е. сокращаться. В пользу такого механизма говорит, в частности, тот факт, что при мышечном сокращении ширина зоны I и зоны Н уменьшается, тогда как ширина зоны А не меняется. Сейчас эта гипотеза окончательно подтверждена, повсеместно признана и называется теорией скользящих нитей. М 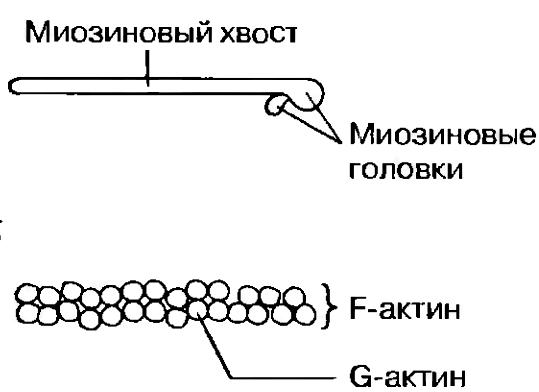 иозиновые (толстые) миофиламенты иозиновые (толстые) миофиламентыМолекула миозина состоит из двух частей: длинного палочкообразного участка («хвоста») и присоединенного к одному из его концов глобулярного участка, который представлен двумя одинаковыми «головками» Молекулы миозина расположены на миозиновой нити таким образом, что головки регулярно распределяются по всей ее длине. В тех местах, где нити актина и миозина перекрываются, миозиновые головки могут прикрепляться к соседним актиновым нитям. Насколько важно такое взаимодействие, мы оценим при рассмотрении собственно механизма сокращения саркомера. Актиновые (тонкие) миофиламенты Каждый актиновый миофиламепт образован двумя цепочками из глобулярных молекул актина (G-актина), закрученными одна вокруг другую наподобие спирали Весь комплекс актиновых молекул называется F-актином (фибриллярным актином). Полагают, что с каждой молекулой G-актина связана одна молекула АТФ. Механизм сокращения С 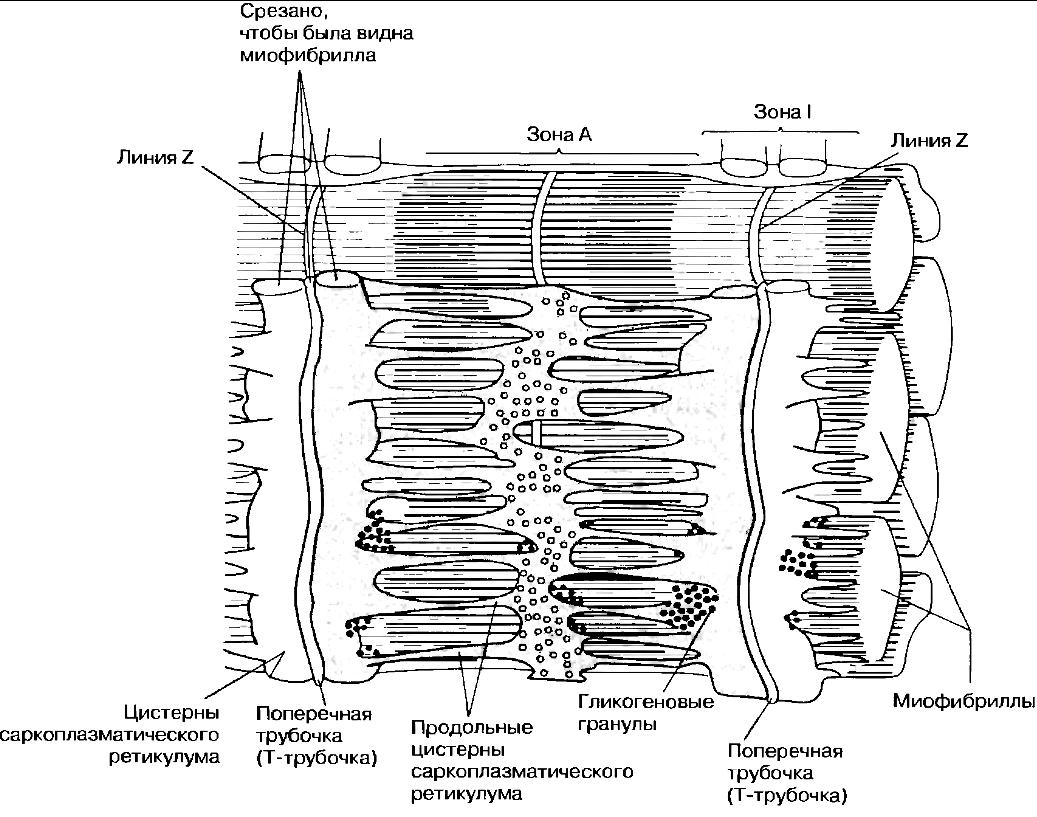 хема механизма сокращения следующая. Там, где актиновые и миозиновые миофиламенты перекрываются, миозиновые головки как крючки «зацепляются» за соседние F-актиновые нити, образуя с ними поперечные МОСТИКИ. Эти МОСТИКИ загибаются, как пальцы, в одном направлении, протаскивая актиновые миофиламенты вдоль миозиновых. Затем головки отделяются от актина, распрямляются, соединяются с новыми его участками, и цикл повторяется. При сокращении в каждый данный момент времени примерно половина головок «тянет», а остальные возвращаются в исходное положение, что обеспечивает плавность процесса. Энергию для него дает АТФ. Молекулы АТФ гидролизуются до АДФ и фосфата под действием АТФазы, содержащейся в миозиновых головках. хема механизма сокращения следующая. Там, где актиновые и миозиновые миофиламенты перекрываются, миозиновые головки как крючки «зацепляются» за соседние F-актиновые нити, образуя с ними поперечные МОСТИКИ. Эти МОСТИКИ загибаются, как пальцы, в одном направлении, протаскивая актиновые миофиламенты вдоль миозиновых. Затем головки отделяются от актина, распрямляются, соединяются с новыми его участками, и цикл повторяется. При сокращении в каждый данный момент времени примерно половина головок «тянет», а остальные возвращаются в исходное положение, что обеспечивает плавность процесса. Энергию для него дает АТФ. Молекулы АТФ гидролизуются до АДФ и фосфата под действием АТФазы, содержащейся в миозиновых головках. Между двумя последовательными образованиями мостиков одной головкой проходит 1/50-1/100 с. Очевидно, для этого необходимо очень интенсивное расходование АТФ. Этим объясняется наличие в мышечном волокне огромного числа митохондрий, которые восполняют запас АТФ в процессе аэробного дыхания. Саркомер способен укоротиться на 30-60% исходной длины. Как запускается и прекращается процесс сокращения? Его активируют ионы кальция (см. ниже «Роль тропомиозина и тропонина»). Они накапливаются в саркоплазматическом ретикулуме (специализированном эндоплазматическом ретикулуме мышечного волокна), который образует расширенные цистерны вокруг линий Z. Эти цистерны контактируют с поперечными трубочками (Т-трубочками, или Т-системой), образованными впячиванием сарколеммы и пронизывающими саркоплазму (цитоплазму мышечного волокна). Когда нервный импульс по мотонейрону достигает нервно-мышечного соединения на поверхности мышечного волокна, потенциал действия из области концевой пластики волной распространяется по Т-системе, передается от нее саркоплазматическому ретикулуму и приводит к высвобождению ионов кальция в саркоплазму. Роль тропомиозина и тропонина Актиновые филаменты состоят из F-актина и двух вспомогательных белков — тропомиозина и тропонина. Молекулы тропомиозина образуют две довольно растянутые спиральные цепочки, которые обвивают F-актин. Тропомиозин служит для включения и выключении механизма сокращения. С тропомиозином связан глобулярный белок тропонин, способный обратимо присоединять ионы кальция. Когда мышца в покое (расслаблена), тропомиозин блокирует на тонком миофиламенте участки для прикрепления миозиновых головок, «отключая» актин. Ионы кальция, высвобождаясь из саркоплазматического ретикулума, соединяются с тропонином. заставляя его и одновременно тропомиозин, с которым он связан, сдвинуться. Это приводит к разблокированию участков прикрепления миозиновых головок — актин «включается», и начинается скольжение нитей по описанному выше механизму. Когда раздражение мышечного волокна стимулирующими импульсами прекращается, ионы кальция закачиваются из саркоплазмы в саркоплазматический ретикулум кальциевым насосом в ее мембране, т. е. путем активного транспорта, также требующим энергии АТФ. Тропонин и тропомиозин приобретают исходную конфигурацию, тонкий миофиламент «отключается», и мышечное волокно расслабляется. Источники энергии. Источниками энергии для мышечного сокращения обычно служит глюкоза, приносимая кровью или образующаяся при расщеплении гликогена в мышцах, а также жирные кислоты. При окислении этих молекул в митохондриях (аэробном дыхании) синтезируется АТФ. Обычно кислород для дыхания поставляется гемоглобином крови. Однако мышцы могут также запасать его, поскольку содержат белок миоглобин, близкий по структуре к гемоглобину. Миоглобин также обратимо связывается с кислородом (оксигенируется) и высвобождает ею в случае необходимости, когда кровь не успевает удовлетворять потребности мышечной ткани в кислороде, например при интенсивной физической нагрузке. В расслабленной мышце уровень АТФ низок, поэтому АТФ быстро расходуется при сокращении и запас должен пополняться за счет иных механизмов, пока скорость аэробного дыхания не адаптируется к возросшим энергозатратам. Один из способов регенерации АТФ в анаэробных условиях основан на использовании креатинфосфата. Это вещество всегда присутствует в мышце, но его запасов обычно хватает ненадолго — за 1 мин интенсивной физической работы расходуется примерно 70% креатинфосфата. Следовательно, креатинфосфат полезен лишь в случае кратковременной и интенсивной мышечной активности, например при резком рывке во время спринтерского бега. Затем его запасы должны пополняться за счет окисления жирных кислот или глюкозы. При интенсивной работе мыши кислород быстро расходуется и аэробное дыхание становится невозможным. В таких условиях мышцы регенерируют АТФ за счет анаэробного расщеплении глюкозы. В этом случае говорят, что работа мышцы создает кислородную задолжность. Одним из конечных продуктов анаэробного получения АТФ является молочная кислота. Накапливаясь в мышцах, она изменяет их кислотно-щелочной баланс, что выражается в повышенной утомляемости, боли, а иногда и в спазмах. Время полной переработки молочной кислоты — это именно то время, которое необходимо для ликвидации кислородной задолженности после энергичной работы мыши. (к оглавлению) Путем тренировки можно повысить устойчивость организма к молочной кислоте и, следовательно, увеличить объем развивающейся кислородной задолженности. Медленные и быстрые мышечные волокна Выделяют два типа скелетных мышечных волокон, каждый из которых имеет свои физиологические особенности. Это медленные (тонические) и быстрые (фазические волокна). В некоторых мышцах могут быть только быстрые или только медленные волокна, в других — волокна обоих типов в определенном соотношении. Благодаря волокнам этих двух типов организм способен передвигаться и поддерживать позу. Быстрые волокна позволяют мышце сокращаться с высокой скоростью. В большом количестве эти волокна имеются у хищников; они обеспечиваю! быстроту реакций при ловле добычи. Вместе с тем потенциальная добыча, чтобы не стать жертвой хищников, тоже должна быть способна к быстрому реагированию. В обоих случаях от подвижности животного будут зависеть его шансы на выживание. Когда животное находится в покое, оно поддерживает определенную позу с помощью тонических мышечных волокон. Им свойственно более медленное и длительное сокращение, но зато энергетические затраты при этом меньше, чем при сокращении быстрых волокон. У человека все мышцы тела состоят из волокон обоих типов, но обычно один из них доминирует. Это имеет физиологическое значение, поскольку тонические мышцы способны к медленному и длительному сокращению и их соответственно больше в позных мышцах-разгибателях, тогда как сгибателях, предназначенных для быстрых реакций, преобладают фазические волокна. Быстрые мышечные волокна иногда называют белыми: в них относительно мало красного пигмента миоглобина, связывающего кислород. В медленных волокнах его намного больше и их называют красными.
Влияние тренировки на работоспособность мышц Хотя в принципе, объем мускулатуры индивидуума определяется генетически, его можно увеличить путем физических упражнений еще примерно на 60%. Это происходит главным образом за счет увеличения диаметра мышечных волокон и в меньшей степени - числа их самих и миофибрилл. Долговременные биохимические изменения При регулярной тренировке растут число и размеры митохондрий внутри мышечных волокон. В результате процессы, связанные с аэробным дыханием — цикл Кребса, электронный транспорт и окисление жирных кислот — протекают более интенсивно. Способность митохондрий генерировать АТФ может увеличиться вдвое. В волокнах накапливается больше креатинфосфата, гликогена, жира, а также миоглобина. При обогащении мышц последним растут запасы кислорода. В совокупности это приводит к тому, что у спортсмена при физической нагрузке уменьшается зависимость от анаэробного дыхания, и. следовательно, в мышцах накапливается меньше лактата. Вместе с тем растет способность организма мобилизировать в энергетических целях запасы липидов, расщепляя их с образованием жирных кислот. Следовательно, спортсмену легче, чем нетренированному человеку «сбросить вес» путем физических упражнений. Долговременное увеличение мышечной силы Развиваемая мышцей сила увеличивается только в том случае, если она работает с большей, чем раньше, нагрузкой. (к оглавлению) Это происходит при увеличении как интенсивности работы, так и ее общей продолжительности. Мышцы, вынужденные развивать близкую к предельной для себя силу сокращения, становятся сильнее очень быстро, даже если тренировки занимают ежедневно по несколько минут. Однако для сохранения этого результата тренировки должны быть регулярными. В противном случае мышцы возвращаются к исходному состоянию, утрачивают силу и способность часто сокращаться, и спортсмен, как говорится, теряет форму. Кровоснабжение мышц при регулярных тренировках увеличивается число сосудов, снабжающих мышцы кровью. Это создает более эффективную систему обеспечения мышц кислородом и глюкозой, а также удаления конечных продуктов обмена. При длительных тренировках кровеносная и дыхательная системы адаптируются таким образом, что появляющаяся после первых упражнений кислородная задолжность в дальнейшем может быть полностью компенсирована. Способность мышц к продолжительной интенсивной работе обычно зависит от скорости и эффективности поглощения и использования ими кислорода. Координация Тренировка улучшает координацию работы мышц, составляющих антагонистические пары, что позволяет совершать более сложные и точные движения; повышается также скорость сокращения и расслабления. Последнее особенно важно, поскольку, если мышца не успевает вовремя расслабиться, то она может быть разорвана тянущим усилием антагониста. Растяжение и разрыв мышц может произойти под действием чрезмерной нагрузки. Чтобы снизить вероятность травм, применяют специальные «согревающие» упражнения. После сильных нагрузок в мышцах обычно сохраняется некоторое остаточное напряжение. Чтобы избавиться от него, используют расслабляющие упражнения, в ходе которых осторожно растягивают мускулатуру. Гладкие мышцы Гладкая мускулатура имеется в стенках многих полых образований в теле животного, к которым относятся кишечный тракт, мочевой пузырь, кровеносные сосуды, мочеточники, матка, семявыносящий проток. Гладкомышечные клетки - одноядерные, веретенообразные; они скреплены соединительной тканью, состоящей в основном из коллагена. Клетки располагаются параллельно друг другу и образуют отдельные мышечные слои. Это можно видеть на примере гладкой, мускулатуры кишечника, где имеются наружный продольный и внутренний кольцевой слои. Когда сокращается продольный слой, кишка укорачивается и ее просвет расширяется, в то время как сокращение кольцевого слоя приводит к удлинению кишки и сужению просвета. Такая координированная работа мышц, называемая перистальтикой, способствует перемещению содержимого кишки; это хороший пример важной функции гладкой мускулатуры, а именно передвижения веществ внутри полых органов. О 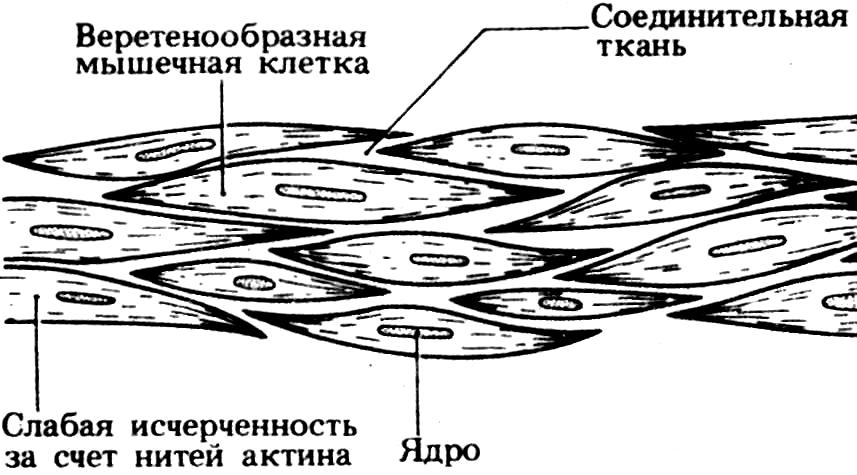 тдельная гладкомышечная клетка в расслабленном состоянии имеет около 50-200 мкм в длину и 2-5 мкм в диаметре. Актин расположен в ней продольными тяжами. Сейчас считается общепризнанным, что гладкие мышцы позвоночных обычно содержат и миозиновые нити, которые, однако, могут отличаться от миозиновых нитей поперечнополосатых мышечных волокон. Поперечной исчерченности здесь нет, так как актиновые и миозиновые нити не расположены строго упорядоченным образом. Как полагают, механизм сокращения гладких и поперечнополосатых мышц в основе своей одинаков, хотя регуляция их работы может быть совершенно различной. тдельная гладкомышечная клетка в расслабленном состоянии имеет около 50-200 мкм в длину и 2-5 мкм в диаметре. Актин расположен в ней продольными тяжами. Сейчас считается общепризнанным, что гладкие мышцы позвоночных обычно содержат и миозиновые нити, которые, однако, могут отличаться от миозиновых нитей поперечнополосатых мышечных волокон. Поперечной исчерченности здесь нет, так как актиновые и миозиновые нити не расположены строго упорядоченным образом. Как полагают, механизм сокращения гладких и поперечнополосатых мышц в основе своей одинаков, хотя регуляция их работы может быть совершенно различной.Возбуждение в гладкой мускулатуре распространяется относительно медленно, что обусловливает медленное длительное сокращение мышцы и столь же длительный период расслабления. Мышцы способны также к самопроизвольным ритмическим сокращениям, которые могут быть разной частоты и силы. Растяжение гладкой мускулатуры полого органа при наполнении его содержимым обычно сразу же ведет к, ее сокращению, и таким образом обеспечивается проталкивание содержимого дальше. Клетки иннервируются не соматической нервной системой, а двумя видами вегетативных нервных волокон: одни из них относятся к симпатической, а другие - к парасимпатической нервной системе. Противоположное действие, которое оказывают эти волокна на иннервируемый ими орган, позволяет быстро изменять состояние органа в соответствии с возможными изменениями условий. На активность гладких мышц могут также влиять адреналин и ряд других гормонов. Строение сердечной мышцы С 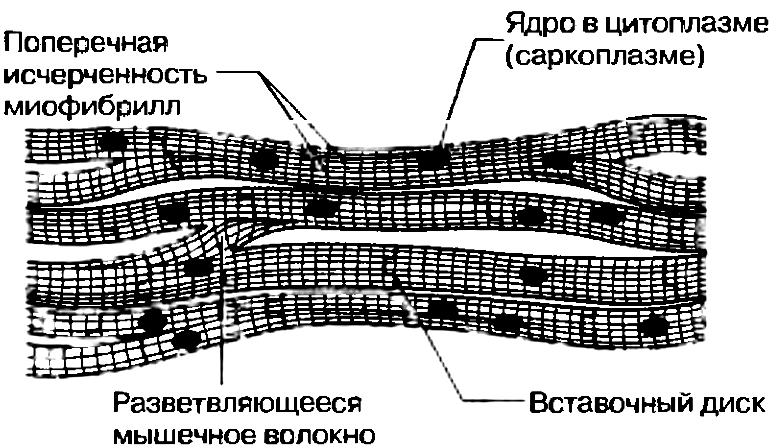 тенка сердца образована сердечными мышечными волокнами, соединительной тканью и мелкими кровеносными сосудами. Каждое мышечное волокно (кардиомиоцит) содержит одно или два ядра, множество крупных митохондрий и множество параллельных друг другу миофибрилл. Миофибриллы образованы актиновыми и миозиновыми нитями (миофиламентами), которые обеспечивают сокращение кардиомиоцита подобно тому, как это происходит в скелетной мышце. В принципе внутреннее строение кардиомиоцитов такое же, как у волокон скелетных мышц, поэтому под микроскопом они также выглядят поперечно-полосатыми. Темные полосы, называемые интеркалярными или вставочными дисками, представляют собой поверхностные клеточные мембраны, отделяющие одну мышечную клетку от другой. Мембраны модифицированы, что позволяет ионам быстро диффундировать сквозь них. Это в свою очередь обеспечивает быстрое распространение возбуждения (потенциала действия) по всей сердечной мышце. Поскольку эти клетки соединены друг с другом и образуют сложную сеть, возбуждение, возникающее в одной из них, тут же охватывает весь миокард, который в результате действует как единое целое. Такая особенность объясняет отсутствие в стенке сердца управляющих его работой нейронов. Сокращается миокард медленнее, чем скелетные мышцы, и утомляется не так быстро. тенка сердца образована сердечными мышечными волокнами, соединительной тканью и мелкими кровеносными сосудами. Каждое мышечное волокно (кардиомиоцит) содержит одно или два ядра, множество крупных митохондрий и множество параллельных друг другу миофибрилл. Миофибриллы образованы актиновыми и миозиновыми нитями (миофиламентами), которые обеспечивают сокращение кардиомиоцита подобно тому, как это происходит в скелетной мышце. В принципе внутреннее строение кардиомиоцитов такое же, как у волокон скелетных мышц, поэтому под микроскопом они также выглядят поперечно-полосатыми. Темные полосы, называемые интеркалярными или вставочными дисками, представляют собой поверхностные клеточные мембраны, отделяющие одну мышечную клетку от другой. Мембраны модифицированы, что позволяет ионам быстро диффундировать сквозь них. Это в свою очередь обеспечивает быстрое распространение возбуждения (потенциала действия) по всей сердечной мышце. Поскольку эти клетки соединены друг с другом и образуют сложную сеть, возбуждение, возникающее в одной из них, тут же охватывает весь миокард, который в результате действует как единое целое. Такая особенность объясняет отсутствие в стенке сердца управляющих его работой нейронов. Сокращается миокард медленнее, чем скелетные мышцы, и утомляется не так быстро.(к оглавлению) |