Глюкагон и адреналин. реферат. Глюкагон и адреналин. Их роль в регуляции углеводного и липидного обмена
 Скачать 1.6 Mb. Скачать 1.6 Mb.
|

 Реферат На тему: Глюкагон и адреналин. Их роль в регуляции углеводного и липидного обмена.  Глюкагон Строение Представляет собой полипептид, включающий 29 аминокислот с молекулярной массой 3,5 кДа и периодом полураспада 3-6 мин. Синтез Осуществляется в клетках поджелудочной железы и в клетках тонкого кишечника. Регуляция синтеза и секреции Уменьшается количество гормона при повышении концентрации глюкозы в крови. Механизм действия Аденилатциклазный. Мишени и эффекты Конечным эффектом является повышение концентрации глюкозы и жирных кислот в крови. Жировая ткань повышает активность внутриклеточной гормон-чувствительной ТАГ-липазы и, соответственно, стимулирует липолиз. Печень активация глюконеогенеза и гликогенолиза, за счет повышенного поступления жирных кислот из жировой ткани усиливает кетогенез. Катехоламины Строение Гормоны надпочечников адреналин и норадреналин под общим названием катехоламины представляют собой производные аминокислоты тирозина. Роль адреналина является гормональной, норадреналин преимущественно является нейромедиатором. Синтез Осуществляется в клетках мозгового слоя надпочечников (80% всего адреналина), синтез норадреналина (80%) происходит также в нервных синапсах. 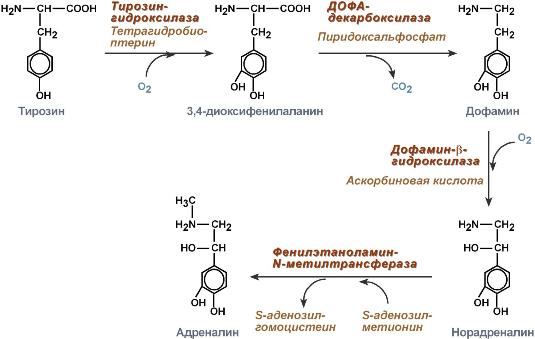 Реакции синтеза катехоламинов Регуляция синтеза и секреции Активируют: стимуляция чревного нерва, стресс. Уменьшают: гормоны щитовидной железы. Механизм действия Механизм действия гормонов разный в зависимости от рецептора. Степень активности рецептора ожет изменяться от концентрации соответствующего лиганда. Например, в жировой ткани при низких концентрациях адреналина более активны α2-адренорецепторы, при повышенных концентрациях (стресс) – стимулируются β1-, β2-, β3-адренорецепторы. Кальций-фосфолипидный механизм при возбуждении α1-адренорецепторов. Аденилатциклазный механизм при задействовании α2-адренорецепторов аденилатциклаза ингибируется, при задействовании β1- и β2-адренорецепторов аденилатциклаза активируется. Мишени и эффекты α1-Адренорецепторы При возбуждении α1-адренорецепторов происходит: 1. Активация гликогенолиза и глюконеогенеза в печени. 2. Сокращение гладких мышц кровеносных сосудов в разных областях тела, мочеточников и сфинтера мочевого пузыря, предстательной железы и беременной матки, радиальной мышцы радужной оболочки, поднимающих волос, капсулы селезенки. 3. Расслабление гладких мышц ЖКТ и сокращение его сфинктеров, α2-Адренорецепторы При возбуждении α2-адренорецепторов происходит: снижение липолиза в результате уменьшения стимуляции ТАГ-липазы, подавление секреции инсулина и секреции ренина, спазм кровеносных сосудов в разных областях тела, расслабление гладких мышц кишечника, стимуляция агрегации тромбоцитов. β1-Адренорецепторы Возбуждение β1-адренорецепторов (есть во всех тканях) проявляется в основном: активация липолиза, расслабление гладких мышц трахеи и бронхов, расслабление гладких мышц ЖКТ, увеличение силы и частоты сокращений миокарда (ино- и хронотропный эффект). β2-Адренорецепторы Возбуждение β2-адренорецепторов (есть во всех тканях) проявляется главным образом: 1. Стимуляция гликогенолиза и глюконеогенеза в печени, гликогенолиза в скелетных мышцах, 2. Усиление секреции инсулина, тиреоидных гормонов. 3. Расслабление гладких мышц трахеи и бронхов, желудочно-кишечного тракта, беременной и небеременной матки, кровеносных сосудов в разных областях тела, мочеполовой системы, капсулы селезенки, 4. Усиление сократительной активности скелетных мышц (тремор), 5. Подавление выхода гистамина из тучных клеток. В целом катехоламины отвечают за биохимические реакции адаптации к острым стрессам, связанным с мышечной активностью – "борьба или бегство": усиление продукции жирных кислот в жировой ткани для работы мышц, мобилизация глюкозы из печени для повышения устойчивости ЦНС, поддержание энергетических потребностей работающих мышц за счет поступающей глюкозы и жирных кислот, снижение анаболических процессов через уменьшение секреции инсулина. Адаптация также прослеживается в физиологических реакциях: мозг – усиление кровотока и стимуляция обмена глюкозы, мышцы – усиление сократимости, сердечно-сосудистая система – увеличение силы и частоты сокращений миокарда, увеличение артериального давления, легкие – расширение бронхов, улучшение вентиляции и потребления кислорода, кожа – снижение кровотока, ЖКТ и почки – снижение деятельности органов, не помогающих задаче срочного выживания. Регуляция углеводного обмена Гормоны. К гормонам, которые влияют на углеводный обмен, принадлежат пептиды инсулин и глюкагон, глюкокортикоид кортизол и катехоламин адреналин. Инсулин индуцирует синтез de novo гликоген-синтазы [1], а также некоторых ферментов гликолиза [3, 5, 7]. Одновременно инсулин подавляет синтез ключевых ферментов глюконеогенеза (репрессия, [4, 6, 8, 9]). Глюкагон как антагонист инсулина действует в противоположном направлении: индуцирует ферменты глюконеогенеза [4, 6, 8, 9] и репрессирует пируваткиназу [7], ключевой фермент гликолиза. Другие аффекты глюкагона основаны на взаимопревращении ферментов и опосредованы вторичным мессенджером цАМФ. По этому механизму тормозится синтез гликогена [1] и активируется расщепление гликогена [2]. Подобным образом действует и адреналин. Торможение пируваткиназы [7] глюкагоном также обусловлено взаимопревращением ферментов. Глюкокортикоиды, прежде всего кортизол индуцируют все ключевые ферменты глюконеогенеза [4, 6, 8, 9]. Одновременно они индуцируют ферменты деградации аминокислот и обеспечивают тем самым глюконеогенез исходными соединениями. Фруктозо-2,6-дифосфат Важную роль в обмене веществ в печени играет фруктозо-2,6-дифосфат. Это сигнальное вещество образуется в незначительных количествах из фруктозо-6-фосфата и выполняет чисто регуляторную функцию: стимулирует гликолиз путем активации фосфофруктокиназы и подавляет глюконеогенез с помощью торможения фруктозо-1,5-дифосфатазы. Образование и распад фруктозо-2,6-дифосфата катализируются одним и тем же белком [10а и б]. В нефосфорилированной форме этот белок вызывает образование фруктозо-2,6-дифосфата [10а]. После фосфорилирования цАМФ-зависимой киназой он действует как фосфатаза [10б] и катализирует превращение фруктозо-2,6-дифосфата в фруктозо-6-фосфат. В присутствии адреналина и глюкагона в клетках печени повышается уровень цАМФ, т.е. оба гормона воздействуют как на гликолиз, так и на глюконеогенез. Суммарным результатом является быстрое повышение уровня глюкозы в крови.  Обмен жирных кислот и ТАГ регулируется Гормональная регуляция Синтез и окисление триацилглицеролов и жирных кислот зависит от соотношения инсулин / глюкагон. 1. Изменение количества ферментов Ферменты комплекса пальмитатсинтазы и ацетил-SКоА-карбоксилазы являются адаптивными ферментами, количество их возрастает при усиленном питании и уменьшается при голодании и потреблении жира. Индуктором биосинтеза этих ферментов является инсулин. 2. Ковалентная модификация Благодаря инсулину, глюкагону, адреналину, тиреотропному и адренокортикотропному гормонам происходит ковалентная модификация ферментов ацетил-SКоА-карбоксилазы и ТАГ-липазы путем фосфорилирования-дефосфорилирования. 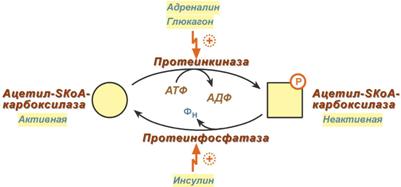 Инсулин активирует протеинфосфатазу и способствует дефосфорилированию и активации ацетил-SКоА-карбоксилазы. Одновременно в клетке дефосфорилируется и инактивируется ТАГ-липаза. Глюкагон, адреналин или другие гормоны, действуя по аденилатциклазному механизму с участием цАМФ-зависимой протеинкиназы, вызывают фосфорилирование и ингибирование ацетил-SКоА-карбоксилазы и, следовательно, останавливают липогенез. Одновременно они активируют ТАГ-липазу (механизм). При уменьшении количества инсулина и возрастании глюкагона усиливаются липолиз в жировой ткани, поступление жирных кислот в печень и другие ткани и реакции их β-окисления. Такое состояние наблюдается при гипогликемии любого происхождения. При обратном соотношении гормонов начинаются реакции синтеза жиров.  |