Синапсы. Httpsmedfsh ruteoriyateoriyaponormalnoyfiziologiilektsiiponormalnojfiziologiisinapsfiziologiyamyshechnyhvolokon Физиология химических синапсов что такое синапс, его определение, виды синапсов, механизмы функционирования химических синапсов
 Скачать 1.08 Mb. Скачать 1.08 Mb.
|

|
Физиология скелетных мышц. Известно три вида мышц: 1) исчерченные (поперечно-полосатые) мышцы; 2) сердечная мышца; 3) неисчерченные (гладкие) мышцы. Поперечно-полосатые скелетные мышцы и их значение, строение и физиологические свойства Скелетные мышцы у человека занимают примерно 40-50% от массы тела. По разным данным, в нашем организме их насчитывается от 400 до 500. Это произвольные мышцы, т.е. они не могут сокращаться без «приказа» из ЦНС. Другими словами, они не обладают автоматизмом. Значение скелетных мышц: 1) поддержание позы человека в пространстве; 2) перемещение тела в пространстве; 3) перемещение частей тела относительно друг друга; 4) обеспечение дыхательной функции; 5) выработка тепла; 6) помощь движению крови и лимфы; 7) участие в осуществлении половых функций; 8) механическая защита внутренних органов; 9) депонирование воды и соли; 10) участие в работе произвольных сфинктеров; 11) участие в осуществлении витальных рефлексов. О значении скелетных мышц красиво сказал И.М.Сеченов: «Смеётся ли ребёнок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к Родине, дрожит ли девушка при первой мысли о любви, создаёт ли Ньютон мировые законы и пишет их на бумаге – везде и всюду окончательным актом является мышечное движение». Строение и физиологические свойства скелетных мышц. Морфологически скелетные мышцы состоят из мышечных волокон (fibra, лат. – волокно) длиной до 3 см и диаметром от 10 до 100 мкм, т.е. представляют собой надклеточную структуру – симпласт – это участок протоплазмы, ограниченный плазмолеммой и содержащий большое количество ядер. Симпласты образуются путём слияния клеток, в данном случае миосимпласт (поперечно-полосатое мышечное волокно) образуется в эмбриогенезе путём слияния клеток миобластов. В каждом волокне содержится до 1000 и более мелких волокон – миофибрилл, имеющих диаметр 1-3 мкм. В каждой миофибрилле содержится 2500 миофиламентов (протофибрилл), которые представляют собой полимеризованные молекулы белков: актина и миозина. Электронно-микроскопическую структуру миофибрилл разберём позже. Физиологические свойства скелетных мышц: 1) возбудимость – способность генерировать потенциал действия; 2) проводимость – способность проводить волну возбуждения; 3) сократимость – способность укорачиваться или развивать напряжение; 4) эластичность – способность развивать напряжение при растягивании. _______________ 1Витальные рефлексы. По П.В.Симонову – это пищевой, питьевой, оборонительный, ориентировочный и другие рефлексы, не требующие взаимодействия с другим организмом. Нейромоторные единицы Физиологически скелетные мышцы состоят из нейромоторных единиц (НМЕ). Нейромоторная единица – это структура, состоящая из мотонейрона и комплекса мышечных волокон, который он иннервирует. Аксон мотонейрона, приносящий нервный импульс (потенциал действия), проникает через базальную мембрану и ветвится между ней и плазмолеммой симпласта, участвуя в образовании концевой пластинки (нервно-мышечного синапса). Нервный импульс запускает освобождение в синапсе химических веществ – медиаторов, которые вызывают возникновение потенциала концевой пластинки (локального ответа). Этот локальный ответ является раздражающим фактором для возникновения на плазмолемме симпласта потенциала действия. Каждое мышечное волокно иннервируется самостоятельно. Классификация нейромоторных единиц скелетных мышц представлена на рис.9. 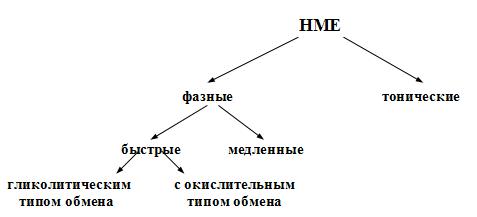 Рис.9. Классификация скелетных нейромоторных единиц мышц. Фазные НМЕ имеют выраженные периоды сокращения и расслабления и делятся на фазные быстрые и фазные медленные НМЕ. Эти НМЕ могут возбуждаться на одиночный импульс раздражения. Фазные быстрые НМЕ имеют мало терминалей и соответственно мало иннервируемых мышечных волокон. В миофибриллах хорошо развит сарко-плазматический ретикулум (СПР), Т-система, а также активен кальциевый насос. Эти НМЕ делятся на НМЕ гликолитического и окислительного типов. У НМЕ гликолитического типа нет миоглобина, поэтому они имеют белый цвет. У них мало митохондрий, АТФ образуется в основном по механизму гликолиза и быстро расщепляется, т.к. в этих НМЕ высокоактивна АТФ-аза. Они сокращаются быстро, сильно и сразу утомляются. Таких НМЕ много в мышцах гортани (НМЕ включает 2-3 миофибриллы) и в мышцах глазного яблока (НМЕ включает 3-6 миофибриллы). Фазные быстрые НМЕ окислительного типа занимают промежуточное положение между фазными быстрыми НМЕ гликолитического типа и фазными медленными НМЕ. В них содержится достаточно много миоглобина, имеется много митохондрий, синтезируется достаточное количество АТФ по механизму окислительного фосфорилирования. Однако активность АТФ-азы у них низкая. Они сокращаются быстро, сильно и достаточно долго. Такие НМЕ преобладают в мышцах длительно летающих птиц. Фазные медленные НМЕ получают иннервацию от -мотонейронов, порог раздражения которых меньше, чем у α-мотонейронов, иннерви-рующих быстрые НМЕ. Аксоны этих нейронов имеют много терминалей и иннервируют соответственно много миофибрилл (от сотен до 2000). У них хуже, чем у быстрых НМЕ, развиты СПР и Т-система. У них много миоглобина, поэтому они имеют красный цвет, много митохондрий, АТФ образуется по механизму окислительного фосфорилирования, а активность АТФ-азы низкая. Эти НМЕ сокращаются медленно, сильно и долго без утомления, а после утомления быстро восстанавливаются. Такие НМЕ преобладают в мышцах, поддерживающих позу человека, четырёхглавой мышце бедра, икроножной и дыхательных мышцах. Тонические НМЕ сокращаются очень медленно слабо и долго (собственно поэтому они называются тоническими). У человека они имеются только в наружных мышцах глаз, но широко представлены в мышцах амфибий и рептилий. Они иннервированы γ-мотонейронами, имеют много терминалей и соответственно образуют синапсы на множестве миофибрилл. На их мембранах нет потенциалзависимых натриевых каналов и поэтому они генерируют только местное возбуждение. Большинство скелетных мышц смешанные, т.е. включают в себя все НМЕ, но в разных соотношениях. Возбудимость разных НМЕ разная. Число тех или других НМЕ зависит от предназначения мышцы. Режимы мышечного сокращения 1. Изометрический режим сокращения (isos, гр. – одинаковый; metron, гр. – мера) – это сокращение, при котором длина мышечного волокна практически не изменяется, а напряжение увеличивается. Размеры саркомеров при этом уменьшаются, а возникающее напряжение приводит к растяжению соединительнотканных (сухожилие, сарколемма) и эластических элементов мышцы, расположенных внутри волокна. Эластическими свойствами обладают продольные L-трубочки (longitudinе, лат. – размер в длину, длительность) СПР, Z-мембраны, поперечные мостики миозиновых филаментов, а также сами актиновые филаменты. В эксперименте изометрическое сокращение можно получить, если изолированную мышцу жёстко закрепить с двух сторон, чтобы она не могла укорачиваться. При этом можно выявить растяжение сухожилий и соединительнотканных элементов мышц, которым передаётся напряжение, развиваемое поперечными мостиками. В атлетической гимнастике упражнения с изометрическим сокращением мышц (например, стремление поднять непосильный груз) вызывают гипертрофию мышц, при этом сила и скорость сокращения мышц увеличиваются. 2. Изотонический режим сокращений (isos, гр. – одинаковый; tonus, гр. – напряжение) – это сокращение, при котором напряжение мышцы практически не изменяется, а меняется её длина. Это все сокращения мышц человека, при которых происходит уменьшение их длины. В атлетической гимнастике упражнения с изотоническим сокращением мышц способствуют развитию их объёма. У человека в чистом виде изотонический и изометрический режимы сокращений не встречаются. Обычно укорочение мышцы сопровождается и развитием напряжения. 3. Ауксотонический режим сокращений (auxos, гр. – и то, и другое; tonus, гр. – напряжение) – это такой режим сокращения, в котором есть элементы изотонического и изометрического сокращения. Типы сокращений Раздражение мышцы или изолированного мышечного волокна одиночным пороговым (надпороговым) стимулом приводит к возникновению одиночного сокращения (рис. 10), которое состоит из трёх периодов: 1) латентного (latentis, лат. – скрытый) периода; 2) периода укорочения (изотонический режим) или напряжения (изометрический режим); 3) периода расслабления. 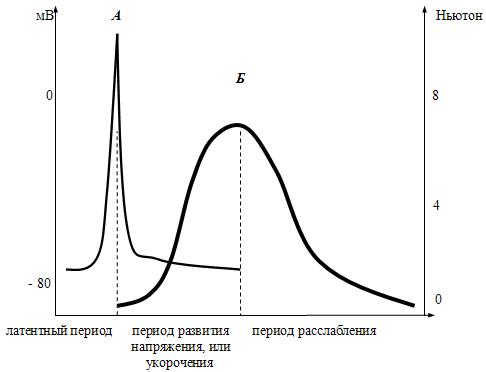 Рис. 10. Периоды одиночного мышечного сокращения: А – потенциал действия; Б – мышечное сокращение. Латентный период (ЛП) – это период от момента нанесения раздражения до начала укорочения мышцы. У лягушки он длится примерно 3 мс. Через 1,5 мс от начала раздражения мышечного волокна возникает латентное расслабление и мышечное волокно удлиняется примерно на 0,001 часть от величены сокращения. В это время увеличивается прозрачность мышечного волокна, уменьшается двойное лучепреломление анизотропного диска, развивается ПД, кальций выходит из СПР, увеличивается pH, уменьшается ёмкостно-омическое сопротивление, выделяется тепло активации и заканчивается абсолютная рефрактерность. С половины латентного периода начинается активация сократительного аппарата – этот переход называется состоянием активности. Период укорочения. При изотоническом режиме сокращения он длится примерно 0,04 секунды (для хладнокровных животных). Сила сокращения, развиваемая мышцей в этот период будет определяться числом нейромоторных единиц, участвующих в сокращении, а сила сокращения каждого волокна будет пропорциональна количеству миофибрилл в нём. Период расслабления. Этот период продолжительнее периода сокращения и длится около 0,05 секунды (у хладнокровных животных), но при утомлении мышцы его длительность значительно увеличивается. В этом периоде происходит уменьшение концентрации кальция в миоплазме и головки миозина отсоединяются от активных центров актиновых филаментов и мышца возвращается в исходное состояние. Сила (амплитуда) одиночного сокращения изолированного мышечного волокна не зависит от силы раздражения, а подчиняется правилу «всё или ничего». В естественных условиях одиночные сокращения скелетных мышц не встречаются. Если в эксперименте нанести на скелетную мышцу дополнительно второй, третий и т.д. раздражающие стимулы в период расслабления, то происходит суммирование возникающих сокращений и результирующее сокращение по силе будет выше, чем сокращение на одиночный стимул – это неполная суммация одиночных сокращений. Картина этих сокращений называется зубчатым тетанусом (tetanus, лат. – суммированный), т.к. на кривой записи видны характерные западения (рис.11А). Тетанус – это сильное и длительное сокращение мышцы. У здорового человека сокращения мышц по типу зубчатого тетануса не встречаются. 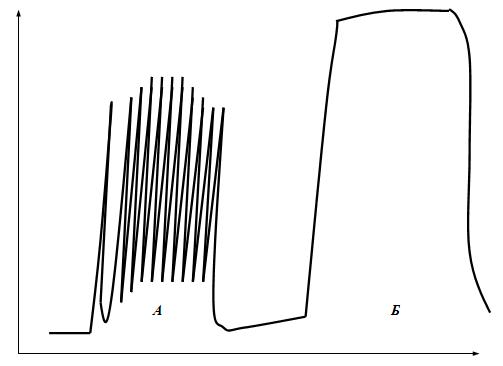 Рис. 11. Зубчатый (А) и гладкий (Б) тетанусы. При увеличении частоты дополнительные стимулы приходятся на период укорочения (или напряжения), и происходит полная суммация одиночных сокращений, которая называется гладким тетанусом (рис.11Б). На кривой записи такого мышечного сокращения нет западений, она гладкая. Амплитуда этой кривой выше, чем у кривых одиночного сокращения и зубчатого тетануса. При этом чем больше частота раздражения (в определённых пределах), тем выше амплитуда тетанического сокращения. Та частота, при которой амплитуда тетануса максимальная, называется оптимальной (optimum, лат. – наилучший). Частоты, которые больше оптимальной, называются пессимальными (pessimum, лат. – наихудший), амплитуда тетануса при этом снижается. В организме к скелетным мышцам в естественных условиях всегда приходит серия импульсов, которые попадают в период укорочения мышцы, т.е. скелетные мышцы человека всегда сокращаются по типу гладкого тетануса. Существует две причины увеличения силы тетанических сокращений в естественных условиях: 1) увеличение числа возбуждённых мотонейронов и синхронизация частоты их возбуждений. Чем больше возбуждённых мотонейронов, тем больше сокращающихся двигательных единиц (это явление пространственной суммации сокращений моторных единиц). Чем больше степень синхронизации мотонейронов, тем больше амплитуда при суперпозиции максимального сокращения, развиваемого каждой двигательной единицы в отдельности; 2) увеличение частоты импульсов, генерируемых каждым мотонейроном (явление временной суммации сокращений каждого волокна данной моторной единицы). Ультраструктурная организация мышечного волокна Чтобы понять механизм мышечного сокращения необходимо знать структурную организацию мышечного волокна. Мышечное волокно имеет длину до 3 см и диаметр от 10 до 100 мкм. В его миоплазме находится до 1000 миофибрилл (диаметр каждой от 1 до 3 мкм), являющихся специализированным сократительным аппаратом. На периферии волокна много ядер, есть митохондрии, хорошо развит СПР, который имеет систему продольных трубочек. Также имеется система Т-трубочек (transversum, лат. – поперечный), представляющая инвагинации (invaginatio, лат. – внедрение, впячивание) плазматической мембраны мышечного волокна, располагающиеся в области Z-мембраны саркомера. Т-трубочки контактируют с концевыми расширениями (цистернами) L-трубочек. Обычно одна Т-трубочка имеет по бокам две цистерны – это называется триада. При световой микроскопии мышечного волокна видна его поперечная исчерченность, т.е. чередование тёмных и светлых участков. При электронной микроскопии оказалось, что исчерченность волокна в поперечном направлении обусловлена особой организацией миофиламентов в миофибриллах. В каждой миофибрилле содержится примерно 2500 миофиламентов – актиновых и миозиновых. Миофиламент (протофибрилла (protos, гр. – первый, первоначальный, первичный)) – это полимеризованные, удлинённые молекулы белков. Актиновый миофиламент представляет собой 400 молекул сократительного белка актина в виде тонких двойных нитей, закрученных в двойную спираль с шагом 36,5 нм. Длина актинового миофиламента составляет примерно 1 мкм, а диаметр – 5 нм. В нём имеются активные центры, располагающиеся друг от друга на расстоянии 20 нм. Молекулярная масса белка актина 42000 дальтон. В бороздках актиновых миофиламентов располагаются молекулы регуляторных белков (эти белки не участвуют прямо в сокращении, но регулируют его) – это тропомиозин и тропонин. Тропомиозин имеет нитевидную форму и к нему прикрепляется тропонин, имеющий глобулярную форму. Тропонин имеет три субъединицы: 1) TN-C – это кальцийсвязывающая субъединица; 2) TN-I – это ингибирующая субъединица, она после связывания TN-C с кальцием изменяет свою конформацию и тропомиозин идёт вглубь (в желобок) спирали, открывая при этом активные центры актинового миофиламента; 3) TN-T – это субъединица, связывающая тропонин с тропомиозином. Миозиновый миофиламент имеет длину примерно 1,6 мкм и диаметр – 10 нм и состоит примерно из 300 молекул белка миозина (молекулярная масса 500000 дальтон). Молекула миозина удлинённая, парная, имеет сдвоенную головку, шейку и хвост. Миозин состоит из двух тяжёлых полипептидных цепей и четырёх лёгких. После обработки миозина трипсином молекула разделяется на быстро седиментирующийся (оседающий) тяжёлый меромиозин (ТММ) и медленно седиментирующийся лёгкий меромиозин (ЛММ). ТММ образует головку и шейку, ЛММ – хвост. ТММ состоит из двух субфрагментов: глобулярного S1, соответствующего головке, и стержневого S2, соответствующего шейке. S1-субфрагмент обладает АТФ-азной активностью и в нём же локализованы центры связывания миозинового филамента с актиновым филаментом и АТФ. Шейка миозина (гибкий участок) представляет собой шарнирное соединение и головка может поворачиваться на шейке вокруг своей оси. Молекулы миозина соединяются между собой хвостами. На боковых сторонах миозинового филамента имеются выступы, которые называются поперечными мостиками, они ориентированы к оси миозиновой нити под углом 120о. Поперечный мостик состоит из головки и шейки миозиновой молекулы. При поляризационной микроскопии мышечного волокна полоски тёмного цвета (вследствие двойного лучепреломления) составляют анизотропный диск А. Он состоит из миозиновых и актиновых филаментов, в его центре имеется светлая полоска Н – это зона, в которой нет актиновых филаментов. В центре Н-полоски имеется М-линия – структура, удерживающая миозиновые филаменты. По обе стороны от диска А видны светлые полоски – это изотропные диски I, обладающие одиночным лучепреломлением поляризованного света. Они образованы только нитями актина. Посередине диска I имеется сетевидная структура, выполняющая опорную функцию – это Z-пластинка. Расстояние от одной Z-пластинки до другой составляет 2,5 мкм и называется саркомером. Саркомер – это функциональная единица сократительного аппарата мышечного волокна. Саркомеры в миофибрилле расположены последовательно и их сокращение вызывает сокращение миофибриллы и общее укорочение мышечного волокна. В покое актиновые и миозиновые филаменты незначительно перекрывают друг друга. На поперечном разрезе миофибриллы видна строго упорядоченная гексогональная организация филаментов: миозиновый филамент окружён шестью актиновыми филаментами. С помощью микроэлектронной техники и интерференционной микроскопии установили, что раздражение плазматической мембраны в области Z-пластинки вызывает сокращение саркомера, при этом длина диска А не меняется, а полоски Н и диск I уменьшаются в размерах. Это говорит о том, что длина актиновых и миозиновых филаментов не меняется, а изменяется область их взаимного перекрытия. На основе этих экспериментальных данных H.Huxley и J.Hanson (1954) выдвинули для объяснения механизма сокращения скелетных мышц теорию скольжения актиновых филаментов относительно миозиновых. |