Синапсы. Httpsmedfsh ruteoriyateoriyaponormalnoyfiziologiilektsiiponormalnojfiziologiisinapsfiziologiyamyshechnyhvolokon Физиология химических синапсов что такое синапс, его определение, виды синапсов, механизмы функционирования химических синапсов
 Скачать 1.08 Mb. Скачать 1.08 Mb.
|

|
Механизм сокращения скелетных мышц Связь между электрическими процессами на плазматической мембране мышечного волокна и сокращением мышцы называется электромеханическим сопряжением и включает в себя процессы электрохимического и хемомеханического преобразования, которые идут последовательно: Электрохимическое преобразование: 1) процесс генерации потенциала действия; 2) распространение потенциала действия вглубь миофибриллы по Т-системе; 3) электрическая стимуляция места контакта мембраны Т-трубочек и цистерн СПР, активация ферментов и повышение концентрации кальция в миоплазме. Хемомеханическое преобразование: 4) взаимодействие кальция с тропонином и деэкранирование активных центров на актиновых миофиламентах; 5) взаимодействие миозиновых головок с актином, вращение головок и развитие тянущего усилия; 6) скольжение актиновых филаментов относительно миозиновых, уменьшение длины саркомеров и укорочение мышечного волокна (или его напряжение). Рассмотрим эти процессы в их последовательности. Потенциал действия с мотонейрона передних рогов спинного мозга по эфферентным нервным волокнам передаётся на плазматическую мембрану мышечного волокна, где терминали этих волокон образуют нервно-мышечные синапсы (концевые пластинки), в которых выделяется медиатор ацетилхолин. В течение длительного времени считалось, что из окончаний каждого нейрона всегда выделяется только один медиатор (принцип Дейла). Однако сейчас доказано, что один и тот же нейрон может выделять два и, возможно, больше медиаторов (сомедиаторов). Так, в концевой пластинке вместе с ацетилхолином выделяется АТФ. Особенности такого совместного действия ацетилхолина и АТФ пока не изучены, но, вероятно, эффект АТФ сводится к определённому типу модуляции передачи возбуждения в синапсе. Ацетилхолин связывается с Н-холинорецептором на постсинаптической мембране, что приводит к возникновению потенциала концевой пластинки (ПКП). ПКП играет роль раздражающего стимула для плазматической мембраны мышечного волокна, на которой возникает потенциал действия, распространяющийся по ней в обе стороны. Далее ПД распространяется по мембране Т-трубочек (мембрана имеет потенциалзависимые натриевые каналы) внутрь мышечного волокна и активирует дигидропиридиновые рецепторы, структурированные в ней. В результате активации рецепторы меняют свою конформационную структуру и активируют рианодинчувствительные кальциевые каналы мембран цистерн L-трубочек СПР. В цистернах ионы Са++ связаны с кальсеквестрином (связывает 45 молекул Са++) и «белком с высоким сродством к кальцию» (связывает 25 молекул Са++). Каналы открываются и кальций по градиенту концентрации диффундирует из цистерн в миоплазму, его концентрация повышается с 10-7 М до 10-5 М. На этом заканчивается электрохимическое преобразование и начинается хемомеханическое. Ионы кальция связываются с TN-C, который имеет ионизированную карбоксильную группу, легко присоединяющую Са++. При этом изменяется конформация TN-C, что приводит к изменению конформации TN-I, в результате чего освобождается место для смещения тропомиозина вглубь образовавшегося желобка между актиновыми миофиламентами. При этом открываются активные (миозинсвязывающие) участки актиновой нити. Головка миозина в покое представляет собой комплекс «миозин + АДФ + фосфат». Активные центры актина обладают большим сродством к этому комплексу, в результате чего происходит присоединение головки миозина к активным центрам актинового филамента. Присоединение актина вызывает быстрое освобождение АДФ и фосфата из миозина, что приводит к изменению конформации головки. При этом головка поворачивается на 45о вокруг своей оси (рабочий ход). Так как головка имеет несколько центров связывания, то они последовательно взаимодействуют с активными участками на актиновом филаменте и при этом развивается тянущее усилие. После поворота головки к ней вместо ушедших АДФ и фосфата присоединяется АТФ, образуя комплекс «миозин + АТФ». Актин обладает к этому комплексу малым сродством, в результате чего происходит отсоединение головки миозина (разрыв поперечных мостиков). Головка становится перпендикулярно к актиновому филаменту. В головке миозина, уже не связанной с актином, происходит гидролиз АТФ, вновь образуется комплекс «миозин + АДФ + фосфат» и головка вновь способна присоединяться к актиновому филаменту. В каждый конкретный момент развития сокращения часть головок миозинового филамента соединена с активными центрами актинового филамента, а другая часть свободна, что позволяет последовательно осуществлять скольжение актиновых нитей вдоль миозиновых, уменьшение длины саркомеров и укорочение мышцы в целом (или развитие напряжения мышцы). В настоящее время появились некоторые факты, которые не вписываются в классическую теорию скольжения: 1) миозиновый филамент при сокращении меняет свою длину и диаметр (утолщается и укорачивается); 2) при сокращении укорачивается не только диск I, но и диск А; 3) между актиновыми и миозиновыми филаментами расстояние – 13 нм, а длина головки миозина – 19-21 нм; 4) сокращение мышцы ступенчатое, а не плавное, как при скольжении. Эти и другие факты позволили Н.С.Мирошниченко и М.Ф.Шуба (1990) высказать гипотезу, что скольжение актиновых филаментов вдоль миозиновых невозможно из-за структурных препятствий и специфического характера действия сил на сократительный аппарат мышцы. Авторы считают, что в основе сокращения лежит вкручивание миозиновых филаментов в трубкообразные структуры, образованные актиновыми филаментами, а само сокращение обеспечивается междоменными перемещениями в работающих по очереди головках миозина. Указанная гипотеза вкручивания в настоящее время находит всё большее и большее признание у специалистов по «молекулярной механике» и может стать новой современной теорией сокращения скелетных мышц в будущем. Энергетика мышечного сокращения Энергия для сокращения мышц образуется при гидролизе АТФ под влиянием фермента АТФ-азы. Свойства этого фермента появляются у головки миозина (тяжёлых цепей), когда она присоединяется к активным центрам актина. В расслабленном состоянии ни миозин, ни актин не обладают АТФ-азной активностью. При расщеплении АТФ меняется конформация АТФ-азного центра головки миозина и головка переходит в новое высокоэнергетическое состояние. Она поворачивается и присоединяется к активному центру актинового филамента. Повторное присоединение головки миозина к актину сопровождается новым поворотом головки. При каждом цикле соединения и разъединения головки миозина с актином гидролизуется одна молекула АТФ на один поперечный мостик. При этом концентрация кальция в миоплазме должна быть не менее 10-5 М. Одна молекула АТФ при расщеплении до АДФ и Н3РО4 выделяет 10000 ккал (48 кДж) энергии. Распад АТФ, т.е. снижение отношения АТФ к АДФ, запускает ресинтез АТФ (принцип Энгельгарта). Ресинтез АТФ осуществляется анаэробными и аэробными путями. Для этого в мышцах имеется три энергетических системы, которые включаются последовательно по мере расходования АТФ: 1) анаэробная фосфагенная система (АТФ-КФ система); 2) гликолитическая система; 3) окислительная система. Эти системы отличаются друг от друга по энергетической ёмкости, т.е. по максимальному образованию энергии в единицу времени. Как только АТФ гидролизовалась до АДФ, мгновенно (через несколько миллисекунд) включается фосфагенная система и происходит срочный анаэробный ресинтез АТФ из креатинфосфата (КФ), т.к. КФ значительно больше, чем АТФ (креатинкиназная реакция): АДФ + КФ → АТФ + К Если истощаются запасы КФ, то АТФ образуется из двух молекул АДФ (миокиназная реакция): 2АДФ → АТФ + АМФ АМФ подвергается дезаминированию: АМФ → ИМФ + NH3. Одновременно идёт ресинтез КФ в митохондриях. Ёмкость фосфагенной системы мала. При её максимальной работе АТФ хватает на 5-6 секунд работы. Если сокращение мышц продолжается, то последовательно развёртываются гликолитическая и окислительная системы. Работу гликолитической системы запускает АДФ. Анаэробно начинают расщепляться глюкоза и гликоген до лактата. При этом одна молекула глюкозы даёт энергию для синтеза двух молекул АТФ. Эта АТФ расходуется на работу мембранных насосов. Образование АТФ анаэробным путём происходит в 2-3 раза быстрее, чем аэробным. Ёмкость гликолитической системы в 25 раз больше, чем фосфагенной, но намного меньше, чем окислительной. Поэтому сокращение мышц при анаэробном гликолизе может быть интенсивным, но будет продолжаться 1-2 минуты, а затем с накоплением молочной кислоты наступает утомление. При продолжающемся сокращении мышц через 2-3 минуты развёртывается окислительная система и ресинтез АТФ будет осуществляться в основном за счёт окислительного фосфорилирования. При этом одна молекула глюкозы даёт 36 молекул АТФ. Ёмкость окислительной системы в тысячи раз превышает ёмкость фосфагенной и гликолитической систем. Поэтому при хорошем кровоснабжении и достаточном поступлении кислорода мышцы работают несколько часов без утомления. Если сокращение мышц длительное, но малоинтенсивное, а потребность мышц в кислороде при этом удовлетворяется полностью, то АТФ ресинтезируется системой окислительного фосфорилирования за счёт окисления жиров. Такая ситуация наблюдается у спортсменов-стайеров (бег на марафонские дистанции). При интенсивном сокращении мышц (выполнение большой работы за короткое время) энергия для сокращения мышц выделяется за счёт окисления углеводов гликолитической системой. Такая ситуация наблюдается во время бега на короткие дистанции у спортсменов-спринтеров. Механизм расслабления и контрактура скелетных мышц В начале расслабление идёт пассивно за счёт эластических компонентов мышцы (сухожилий, связок), а затем активно. Главным моментом в активном расслаблении является секвестрация (sequestrare, лат. – отделять) кальция миоплазмы в его хранилища, т.е. в СПР. Удаление кальция из миоплазмы производит кальциевый насос, главной частью которого является Са++-активируемая Mg++-зависимая АТФ-аза, находящаяся в мембране элементов СПР. В активации этой АТФ-азы принимают участие: 1) большая концентрация кальция в присутствии ионов магния; 2) фосфаты, которые образуются при гидролизе АТФ; 3) белок кальсеквестрин, который находится на внутренней стороне мембраны цистерн СПР и принимает участие в секвестрации. Са++-активируемая АТФ-аза расщепляет АТФ, высвобождается энергия и кальций активно нагнетается против градиента концентрации в цистерны СПР. Концентрация кальция в миоплазме становится равной примерно 10-7 М, головки миозина отсоединяются от активных участков актина, смещается тропомиозин, закрываются активные участки актина, делая его неспособным взаимодействовать с миозином, и мышца расслабляется. Контрактура мышц. Если в мышцах недостаточно АТФ, то энергии для работы кальциевого насоса нет. Концентрация кальция в миоплазме сохраняется высокой, головки миозина не отсоединяются от активных участков актиновых миофиламентов и расслабления не наступает. Такое длительное, иногда необратимое сокращение называется мышечной контрактурой. Она бывает обратимой и необратимой. Контрактура после длительного тетануса (посттетаническая контрактура) является обратимой, т.е. спустя некоторое время она проходит. К необратимым контрактурам относятся тепловая и посмертная (трупное окоченение) контрактуры. Сила и работа скелетных мышц. Коэффициент полезного действия (КПД) Сократительная способность скелетной мышцы характеризуется: 1) силой сокращения; 2) степенью и скоростью развития напряжения; 3) величиной и скоростью укорочения; 4) скоростью расслабления. Сила мышцы – это тот максимальный груз, который мышца в состоянии приподнять (оторвать от земли). Сила мышцы зависит от физиологического поперечного сечения. Физиологическое поперечное сечение мышцы – это сечение, проходящее через все миофибриллы в поперечном направлении независимо от их геометрического расположения. Например, для перистой мышцы (икроножной) оно выглядит следующим образом (рис.12). Чем больше физиологическое поперечное сечение мышцы, тем больше общая сила мышцы. Выделяют также абсолютную силу мышцы – это частное от деления силы мышцы на 1 см2 её площади. Сила мышц определяется динамометрами различной конструкции (кистевой, становой и др.) и выражается в килограммах или граммах. 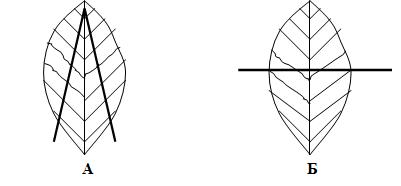 Рис.12. Физиологическое (А) и геометрическое (Б) поперечные сечения перистой мышцы. Зависимость силы сокращения изолированной мышцы от длины саркомера. Сила сокращения изолированной мышцы при прочих равных условиях зависит от исходной длины мышцы. Небольшое растяжение мышцы приводит к увеличению силы сокращения из-за суммирования пассивного напряжения, обусловленного эластическими компонентами мышцы и активного сокращения. Максимальная сила развивается при длине саркомера равной от 2 до 2,2 мкм, т.к. именно при этой длине образуется наибольшее количество актомиозиновых мостиков, развивающих тянущее усилие. Увеличение длины саркомера ведёт к уменьшению силы сокращения, т.к. при этом уменьшается область взаимодействия актиновых и миозиновых нитей (рис.13). Зависимость силы сокращения от скорости сокращения. Сила сокращения мышцы увеличивается при уменьшении скорости сокращения (рис. 14). Из этой зависимости следует, что при большой скорости сокращения мышца обладает малой силой и может переместить небольшой груз, а при снижении скорости сокращения величина перемещаемого груза увеличивается. 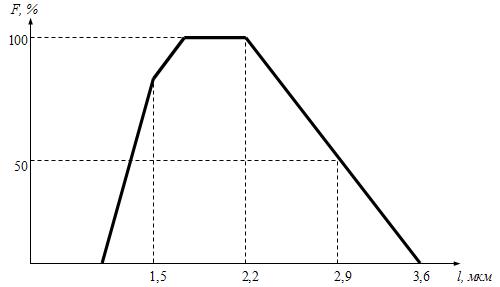 Рис. 13. Зависимость силы мышечного сокращения (F) от длины саркомера (l). 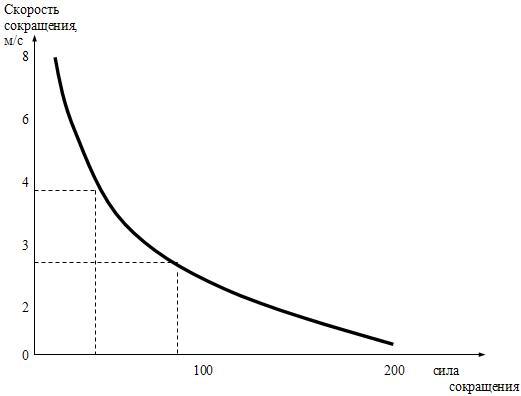 Рис.14. Соотношение между силой и скоростью сокращения мышцы. Работа мышц. Работа мышц определяется произведением её силы на расстояние перемещения груза: A = F•S , где А – работа мышцы; F – сила мышцы; S – расстояние перемещения груза. Если мышца сокращается без нагрузки, значит F = 0, и, следовательно А = 0. Если S = 0, то работа переходит в тепло. Интенсивная аэробная работа мышц лимитируется скоростью потребления кислорода. В этом случае углеводы как субстрат окисления имеют неоспоримое преимущество перед жирами, т.к. для образования одинакового количества АТФ при окислении углеводов затрачивается меньшее количество кислорода. В таком случае особенно эффективно окисление гликогена. Различают работу динамическую и статическую. Динамическая работа – это работа, при которой совершаются движения. Она делится на: 1) преодолевающую, когда момент силы мышцы больше момента силы сопротивления; 2) уступающую, когда момент силы сопротивления больше момента силы мышцы, мышца при этом удлиняется и груз опускается. Более длинные мышцы могут выполнять большую динамическую работу, так как они могут укорачиваться на большую величину и перемещать груз на большее расстояние. Статическая работа – это работа по удержанию груза, когда оба момента силы равны, происходит преимущественно в изометрическом режиме; например, в фиксированной позе. Статическая работа более утомительна чем динамическая. Коэффициент полезного действия. Хемомеханическая реакция в системе актомиозиновых мостиков и все последующие процессы при сокращении мышцы идут с потерей энергии в форме теплоты. Коэффициент полезного действия (КПД) – это коэффициент, показывающий, какое количество от всей затраченной энергии используется для совершения механической работы. Следует заметить, что выделение тепла при этом не бесполезно, так как оно используется на обогрев тела, а мышцы при работе являются основными обогревателями организма. Коэффициент полезного действия мышцы равен частному от деления внешней работы на всю затраченную для выполнения этой работы энергию, выраженному в процентах: 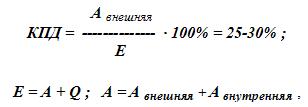 где А внешняя – внешняя работа мышцы; А внутренняя – внутренняя работа мышцы (работа на преодоление сил трения, движения катионов, анионов в мышце); А – общая работа; Е – энергия работы; Q – тепловой выход работы. Отличие работы мышцы от работы технических машин. Человек – это машина, работающая на химической энергии (не тепловой) и поэтому КПД человека выше, чем у машины (на 10-15%). При работе мышцы человека не изнашиваются (как технические устройства), а тренируются. Теплообразование в мышцах Скелетные мышцы являются главными теплообразователями в организме человека. По первому закону термодинамики общая энергия человека и окружающей среды должна быть постоянной. Энергия химических связей в организме превращается в механическую и осмотическую энергию. При этом освобождается тепло. А.Хилл (1922) установил следующие фазы образования тепла. I. Начальное теплообразование, которое делится на 3 вида: 1) тепло активации – это тепло, которое соответствует фазе напряжения сухожилия, выделяется в латентный период сокращения мышц и связано с генерацией потенциала действия, выходом кальция из СПР, соединением кальция с тропонином и работой Na-К-насоса; 2) тепло укорочения – это тепло, которое выделяется в период сокращения мышцы (при изотоническом режиме); 3) тепло расслабления – это тепло, которое выделяется в период расслабления мышцы из её упругих элементов, а также связано с работой кальциевого насоса СПР. Выделение этого тепла связано с гликолитическими процессами в мышце, но не имеет никакого отношения к процессам окислительного фосфорилирования. II. Запаздывающее теплообразование происходит во время восстановительного периода после расслабления мышцы и связано с ресинтезом АТФ в митохондриях путём окислительного фосфорилирования. |