ыв. Метод.реком. к лек_конспект_8 нед (1). Казахский национальный университет им. Альфараби факультет биологии и биотехнологии
 Скачать 1.77 Mb. Скачать 1.77 Mb.
|

РАЗМНОЖЕНИЕ ВОДОРОСЛЕЙВегетативное: диффузное, фрагментация, распад колонии на дочерние; Бесполое (споровое): зооспорами, апланоспорами, автоспорами, эндо-, экзоспорами. Половое: гаметогамия (изогамия, гетерогамия и оогамия); конъюгация. Жизненные циклы развития водорослей. Изоморфный и гетероморфный циклы развития у гаплодиплобионтов. Гаплофазный цикл у гаплобионтов, диплофазный цикл у диплобионтов. Чередование двух поколений (спорофита (2n) и гаметофита (n). Типы редукционного деления в жизненном цикле водорослей (спорическая, гаметическаяи зиготическая). ЭКОЛОГИЯВодоросли водных местообитаний (планктонные, нейстонныеи бентосные). Водоросли вневодных местообитаний (эпилиты,эпипелиты,эндолиты,эндофиты,эпифиты,паразиты,перифитон). Водоросли экстремальных температур (термофильные и криофильныеводоросли). ПРЕДСТАВИТЕЛИ ОСНОВНЫХ ТАКСОНОВОтдел Цианофиты (синезеленые водоросли, цианобактерии): анабена,осциллатория, микроцистис, глеокапса; Отдел Хлорофита (зеленые водоросли, равножгутиковые): хламидомонас,вольвокс,спирогира и др. Отдел Охрофита включает в себя 4 класса: Классзолотистые(динобриони др.) Классжелтозеленые(вошерия, ботридиум и др.) Классдиатомовые (пиннулярия, навикула и диатома и др.) Классбурыеводорослей(ламинария, фукус и др.) Вопросы для самоконтроляДать определение водорослям Водоросли- группа биологическая или систематическая? Рассмотреть эволюцию таллома водорослей На чем основана классификация водорослей? Рассмотреть практическое значение водорослей ЛитератураНестерова, С.Г.. Лабораторный практикум по "систематике растений". - Алматы, 2011 Ботаника. Курс альгологии и микологии.- М., 2007 Лемеза, Н. А. Альгология и микология. Практикум : учеб. пособие / Н. А. Лемеза. – Минск: Выш.школа, 2008. – 319 с. Курс низших растений / редкол. : М. В. Горленко [и др.]. – М. : Высшая школа, 1981. ЛЕКЦИЯ 2. Тема НАСТОЯЩИЕ ГРИБЫЦель лекции: обосновать современную систематику, обсудить структурные и онтогенетические особенности настоящих грибов РО: студент способен проанализировать современную систематику грибов, основываясь на знаниях их морфологии, цитологии, физиологии и геносистематики Вопросы для рассмотрения:Систематика грибов Цитология и морфология грибов; Размножение грибов. Основные представители основных таксонов Термины микологические: грибы, настоящие и псевдогрибы, мицелий, плазмодий, ризомицелий, гаметогамия, гаметангиогамия, соматогамия, хитридиевые, зигомицетовые, сумчатые (аскомицеты), базидиальные и несовершенные грибы, паразиты (биотрофы), некротрофы (сапротрофы), СИСТЕМАТИКА ГРИБОВЦарство грибы (Eumycota, Fungi) Хитридиомикотовых (Chytridiomycota), Зигомикотовых (Zygomycota), Аскомикотовых (Ascomycota) и Базидиомикотовых (Basidiomycota) грибов ЦИТОЛОГИЯ И МОРФОЛОГИЯ ГРИБОВУ настоящих грибов имеются ряд важных признаков: признаки, сближающие их с растениями:образование хорошо выраженной клеточной стенки; осмотрофное питание; размножение спорами; неподвижность в вегетативном состоянии; неограниченный рост; признаки, сближающие грибы с животными:первично гетеротрофный способ питания; наличие в клеточной стенке хитина; запасной продукт – гликоген; выделение продукта жизнедеятельности – мочевины; отсутствие пластид и неспособность к фотосинтезу; специфические признаки:способность к синтезу маннитола (за исключением зигомикотовых грибов), собственные ферментативные системы синтеза триптофана, отсутствие комплекса Гольджи и его замена видоизменениями эндоплазматического ретикулума. Вегетативное тело большинства грибов представляет собой мицелий, состоящий из ветвящихся нитей – гиф с апикальным (верхушечным) ростом и боковым ветвлением. Мицелий пронизывает субстрат и всей поверхностью поглощает из него питательные вещества (субстратныймицелий), а также располагается на его поверхности и может подниматься над субстратом (поверхностный и воздушный мицелий). На воздушном мицелии обычно образуются органы размножения. Различают неклеточный (несептированный, или ценоцитический мицелий) лишенный перегородок, с большим числом ядер, и клеточный (септированный) мицелий, разделенный перегородками – септами на отдельные клетки, содержащие от одного до многих ядер. Для представителей отделов хитридиомикотовых и зигомикотовых характерен неклеточный мицелий. У представителей отделов аскомикотовых и базидиомикотовых мицелий клеточный, с настоящими септами. У дрожжей, вегетативное тело представлено одиночными почкующимися или делящимися клетками, которые не расходятся, образуя псевдомицелий. Некоторые примитивные грибы имеют одноклеточный таллом, лишенный клеточной стенки (ризомицелий– разветвленные нитевидные структуры, лишенные ядер). При формировании плодовых тел и некоторых вегетативных структур гифы грибов плотно переплетаются, образуя ложную ткань – плектенхиму,например, талломы лабульбениомицетов из аскомицетов. У настоящих грибов можно наблюдать видоизменения мицелиев (мицелиальные тяжи, ризоморфы, склероцииидр.),отличающиеся по структуре и выполняемым функциям. У грибов гифы соединяются параллельно в мицелиальные тяжи, выполняющие проводящие функции. Хорошо развитые и дифференцированные мицелиальные тяжи называют ризоморфами.Они известны, например, у опенка осеннего. Ризоморфы достигают нескольких метров длины. Гифы их наружных слоев имеют утолщенные, часто темноокрашенные стенки и выполняют защитную функцию, а внутренние гифы – проводящую. Другой тип видоизменений мицелия представляют широко распространенные у многих групп грибов склероции – плотные переплетения мицелия, служащие для перенесения неблагоприятных условий. Клетки склероциев богаты запасными питательными веществами. Часто склероции дифференцированы на кору – наружные слои клеток, обычно толстостенных и темноокрашенных, и внутреннюю часть, состоящую из тонкостенных светлоокрашенных клеток. Химизм клеточной стенки грибов. Клеточная стенка содержит пигменты(меланины) и хитин. Состав клеточной стенки специфичен для представителей крупных таксономических групп грибов: у хитридиомицетов, аскомицетов и базидиомицетов в клеточных стенках содержатся хитин-глюкановый комплекс; у зигомицетовых грибов клеточные стенки мицелия содержат хитинхитозановыйкомплекс; у дрожжевых грибов клеточные стенки содержат хитин-маннановыйкомплекс. В клетках грибов находится от одного до 20–30 ядер. Их размер обычно около 2–3 мкм. В клетках грибов присутствуют многочисленные гранулы гликогена, капли липидов. В вакуолях часто находятся гранулыбелковиволютина. Зооспорыигаметы(подвижные клетки) встречаются только у хитридиовых грибов.Жгутики у настоящих грибов в отличие от псевдогрибов, бичевидные гладкие, расположены на заднем конце. РАЗМНОЖЕНИЕ ГРИБОВ: вегетативное, бесполое и половое. При вегетативном размножении от мицелия отделяются неспециализированные его части, которые дают начало новому мицелию (хламидоспоры– толстостенные клетки), предназначенных для перенесения неблагоприятных условий. У дрожжей вегетативное размножение происходит путем почкованияклеток. Бесполое размножение происходит при помощи специализированных неполовых клеток– спор. Споры у грибов развиваются эндогенно – внутри спорангиев или экзогенно – на особых гифах мицелия – конидиеносцах. У зигомицетов в спорангиях развиваются неподвижные споры, называемые спорангиоспорами. Спорангии находятся на специализированных гифах – спорангиеносцах, поднимающихся над субстратом. Конидии,как и спорангиоспоры, – неподвижные споры. В отличие от эндогенных спорангиоспор образуются на специализированных ветвях – конидиеносцах экзогенно. Конидии различают по строению - одно-, дву-, многоклеточные; по форме (шаровидные, серповидные, булавовидные, муральные и т.д.); бесцветные, светлоокрашенные (белые, зеленые, желтые, голубые и т.д.) и темноокрашенные (оливковые, черные и темнокоричневые и др.). Конидиеносцы (гифы на которых формируются конидии) бывают одноклеточные и многоклеточные, прямостоячие и разветвленные, часто наблюдается их агрегация в коремии, ложа и пикниды. Бесполое размножение при помощи конидий характерно для базидиомицетов и аскомицетов, в том числе и анаморфных, где бесполое размножение достигло максимального разнообразия. Половое размножение отмечено у всех групп грибов, кроме анаморфных, называемых также несовершенными грибами. Формы полового процесса у грибов разнообразны. Их можно разделить на три большие группы: гаметогамия,гаметангиогамияисоматогамия. ОСНОВНЫЕ ПРЕДСТАВИТЕЛИ ОСНОВНЫХ ТАКСОНОВОсновные представители хитридиевых грибов (отдел Chytridiomycota) Представители отделахитридиомикотовыегрибы(Chytridiomycota) тесно связаны с водной средой, паразитирует на водорослях, других водных грибах, водных высших растениях, на беспозвоночных животных. Некоторые паразиты высших наземных растений, главным образом во влажной почве. Значительно меньшая часть ведет сапротрофный образ жизни. Вегетативное тело представителей этого класса у многих внутриклеточных паразитов в виде голой плазменной массы (плазмодий). Более высокоорганизованные формы имеют ризомицелий или зачаточныймицелий. Основа клеточной стенки хитридиовых грибов – хитин(до 60 %). Бесполоеразмножениеосуществляется зооспорамиразличного строения и формы с одним бичевидным гладким жгутиком, прикрепленным сзади. Полового размножение: хологамия (сливаются сами особи), с помощью гамет (изогамия, гетерогамия и оогамия). Гаметы образуются в специальных клетках органах – гаметангиях. Зигота превращается в покоящуюся клетку – цисту, одетую толстой оболочкой, содержащей хитин. В некоторых случаях цисты образуются без полового процесса. Большинство хитридиевых грибов гаплоидны в вегетативном состоянии, и диплоидная фаза у них представлена зиготой, прорастающей после периода покоя, чаще всего как зооспорангии. Представители: Ольпидиум капустный - возбудитель «черной ножки» капустной рассады. Растения заражаются в парниках, особенно при избыточной влажности почвы. Стебель пораженного растения темнеет, утончается, нередко загнивает, а само растение поникает и гибнет. Синхитриумвнутриклеточный- возбудитель рака картофеля. Основные представители зигомицетовых грибов (отдел Zygomycota)Почти все представители отдела живут на суше. Среди них имеются сапротрофы и паразиты грибов, высших растений, насекомых, других животных и человека. Мицелий сильно ветвится и обычно не имеет перегородок, которые появляются только при старении. Перегородки также отделяют спорангии, оогонии и антеридии. В клеточных стенках мицелия содержатся хитин и хитозан. Бесполое размножение осуществляется неподвижными спорами (спорангиоспоры), развивающимися внутри спорангиев, либо экзогенно на конидиеносцах (конидии). Основная особенность зигомицетовых грибов заключается в половом процессе – зигогамии, при котором сливается содержимое двух клеток- органов полового размножения (оогоний и антеридий),,отделяющихся перегородками от несущих гиф), не дифференцированных на гаметы. Копулирующие клетки большей частью не различимы по величине и внешнему виду, у некоторых же одна из клеток больше, а другая – меньше. На месте слияния клеток, функционирующих у большинства как гаметангии (так как они многоядерны) или как гаметы (в случае их одноядерности), развивается покоящаяся зигоспора. На образовавшейся зиготе (зигоспоре) обычно остаются участки гиф, отделившие гаметангии (суспензоры или зигофоры). Иногда зигота как бы приподнимается на них. Класс Zygomycetesделится на следующие порядки: мукоральные (Mucorales)- плесневые грибы,вызывающие порчу продуктов питании при неправильном хранении, энтомофторальные (Entomophthorales)- возбудители болезней насекомых, зоопагальные (Zoopagales)- возбудители болезней животных. Родумукор (Мисоr) принадлежит центральное место в порядке. Вегетативное тело – в виде несептированного мицелия, пронизывающего субстрат и распростертого по его поверхности. Мицелий белого или серого цвета, спорангии заметные простым глазом в виде буроватых или черных точек. Вопросы для самоконтроляКаковы образ жизни, особенности строения и размножения хитридиевых грибов? Назовите практически значимых представителей хитридиевых грибов. Дайте характеристику отдела зигомикотовые грибы, укажите признаки сходства и отличия данного отдела с уже изученными. Каковы способы размножения и распространения зигомицетовых и хитридиевых грибов? Каков химизм клеточной стенки хитридиевых и зигомицетовых грибов? ЛЕКЦИЯ 3. Тема ДИКАРИОМИЦЕТЫ. АСКОМИЦЕТЫ-ФИТОПАТОГЕНЫЦель лекции: обсудить надотдел Дикариомицеты и его практически значимых представителей РО: студент способен проанализировать сумчатые фитопатогенные грибы, основываясь на знаниях их морфологии, цитологии, физиологии и геносистематики Вопросы для рассмотрения:Общая характеристика аскомицетов; Половой процесс- гаметагамия и аскообразование; Морфология асков (сумок) Фитопатогенные аскомицеты Термины микологические: гаплоидный мицелий, дикарион, диплоидный мицелий; клейстотеций, перитеций и апотеций; гаметангиогамия; аски и аскоспоры; протуникатные и эутуникатные аски; строма; эризифовые грибы; спорыньевые грибы. ОБЩАЯ ХАРАКТЕРИСТИКА АСКОМИЦЕТОВОтдел аскомикотовые грибы (Ascomycota) Отделсумчатые грибы (Ascomycota) один из обширнейших отделов грибов, включающий более 50 000 видов (включая около 16000 видов лишайниковообразующих грибов), с широким диапазоном жизненных форм. сюда относятся дрожжи, представленные одиночными почкующимися клетками, многочисленные микроскопические грибы и грибы с крупными плодовыми телами. Основной признак аскомицетных Г. – формирование в результате полового процесса сумок, или асков, содержащих определенное число эндогенных аскоспор, обычно восемь. Вегетативное тело большинства аскомикомицетов – разветвленный гаплоидный мицелий, состоящий из одноядерных или многоядерных клеток. перегородки (септы) образуются в мицелии. септа развивается центрипетально – от стенок гифы к центру и напоминает сужение диафрагмы в объективе фотоаппарата. в центре септы остается пора, через которую передвигается цитоплазма, а также могут мигрировать органеллы клетки, даже ядра. поры в септах играют существенную роль в переносе питательных веществ по гифам в зону роста. У некоторых аскомикотовых (например, дрожжи) настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися или делящимися клетками, иногда образующими псевдомицелий. Дрожжеподобный рост при определенных условиях наблюдается у некоторых мицелиальных аскомицетов, например, тафриновые, грибов-дерматофитов и др. Основные полисахариды, входящие в состав клеточных стенок аскомикотовых – хитин и глюканы. У дрожжей хитин содержится в небольших количествах. Большую часть полисахаридов клеточной стенки составляют глюканы (до 80–90 %). У дрожжей кроме глюканов обнаружены маннаны – полимеры маннозы. В цикле развития многих аскомикотовых большую роль играет бесполое размножение. Споры бесполого размножения – конидии – образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах разного строения. Конидиеносцы образуются на мицелии одиночно, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных или грушевидных структур с отверстием на вершине (пикниды). Конидиальные спороношения служат для их массового расселения. Для большого числа грибов, встречающихся в природе в гаплоидной конидиальной стадии, совершенные (половые) стадии, телиоморфы, неизвестны. Такие грибы относят к формальному отделу дейтеромикотовые, или несовершенные (анаморфные) грибы. ПОЛОВОЙ ПРОЦЕСС- ГАМЕТАГАМИЯ И АСКООБРАЗОВАНИЕ.Типичный для аскомикотовых половой процесс – слияние двух специализированных клеток мицелия, не дифференцированных на гаметы (гаметангиогамия). Гаметангии разного пола (♀, ♂), которые не формируют гаметы, а сами участвуют в половом процессе (плазмогамия). После их слияния сразу наступает кариогамия, и сумка развивается непосредственно из зиготы. В многоядерных гаметангиях аскомицетов сливаются только два ядра. Зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития таких аскомикотовых имеются только гаплоидная и диплоидная фазы. У более высоко организованных аскомицетов женский гаметангий состоит из двух частей – аскогона и нитевидно вытянутой трихогины, мужской гаметангий – антеридий – одноклеточный. При слиянии содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются, а объединяются попарно, образуя дикарионы. Из аскогона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. Эти гифы ветвятся и разделяются септами на двухъядерные клетки. На аскогенных гифах развиваются сумки. Конечная клетка такой гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, еще одно – в основание. Затем образуются две перегородки, отделяющие по одному ядру в конечную (верхушка крючка) и базальную (основание крючка) клетки. В результате слияния этих клеток восстанавливается дикарион и образуется нижняя клетка, из которой крючок может сформироваться повторно. Средняя двухъядерная клетка крючка развивается в сумку. Сумка увеличивается в размерах, ядра дикариона сливаются, диплоидное ядро делится редукционно и, затем, митотически. В результате формируются аскоспоры в количестве 8, реже 4 или много. В цикле развития высших аскомицетов чередуются три фазы: длительная – гаплоидная(от аскоспор до полового процесса), в течение которой происходит бесполое размножение, непродолжительная – дикарионтическая(аскогенные гифы) и очень короткая – диплоидная(молодая сумка). МОРФОЛОГИЯ АСКОВ (СУМОК)По строению оболочки и функциям сумки делят на две группы: Протуникатныесумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, пассивно освобождая аскоспоры. Эутуникатныесумки характеризуются более плотными, оболочками, часто со специальными приспособлениями для вскрытия сумки. Они активно участвуют в распространении аскоспор. Строение оболочки эутуникатных сумок бывает двух типов: а) оболочка унитуникатных сумок относительно тонкая и выглядит однослойной, на вершине сумок обычно имеется апикальный аппарат различного – строения, служащий для их вскрывания (оперкулятные сумки вскрываются крышечкой, иноперкулятные – щелью и разрывом на вершине); б) битуникатные сумки имеют двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины, внутренний слой под действием повышенного тургорного давления растягивается, и аскоспоры активно выбрасываются. МОРФОЛОГИЯ ПЛОДОВЫХ ТЕЛ АСКОМИЦЕТОВРазличают следующие типы плодовых тел: клейстотеций–замкнутое плодовое тело, сумки расположены беспорядочно или упорядоченным слоем («гимением»), споры высвобождаются пассивно при разрушении 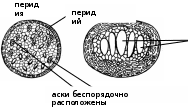 упорядоченный слой асков с аскоспорами упорядоченный слой асков с аскоспорами апотеций – открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен гимений – упорядоченный слой сумок и парафиз апотеций – открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен гимений – упорядоченный слой сумок и парафиз перитеций – полузамкнутое, обычно кувшиновидное плодовое тело с отверстием на вершине, сумки расположены слоем, чередуются со стерильными элементами: парафизами (растут между сумками от основания перитеция), в носике перитеция иногда наблюдаются перифизы – короткие гифы, направленные к выходу; 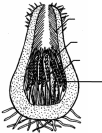 перефизы парафизы перефизы парафизысумки=аски придатки сумчатый гриб рода ФИТОПАТОГЕННЫЕ АСКОМИЦЕТЫЭризифовые или мучнисторосяные (мучнисторосные) грибы- фитопатогенные грибы, возбудители мучнистой росы культурных и дикорастущих растений. Класс леотиомицеты. Порядокэризифальные, или мучнисторосяные (Erysiphales) объединяет аскомицеты, образующие клейстотеции, в которых сумки в зрелости располагаются правильным пучком или слоем. Представители этого порядка – облигатные паразиты высших растений, вызывающие у них заболевания, известные под общим названием «мучнистая роса». Белый, позднее темнеющий мицелий мучнисторосяных грибов находится обычно на поверхности пораженных органов растений. На нем образуются специальные структуры – апрессории, от которых, отходят гаустории, проникающие в клетки эпидермиса растения-хозяина. На мицелии мучнисторосяных через несколько дней после заражения развивается конидиальная стадия – прямые неразветвленные конидиeносцы с цепочками конидий. В это время пораженные органы растения покрыты мучнистым налетом конидий – отсюда название заболевания – «мучнистая роса». Конидии распространяются воздушными течениями и заражают новые растения. Сумчатая стадия развивается у мучнисторосяных в конце периода вегетации. На мицелии гриба образуются аскогоны (без трихогины) и антеридии. Содержимое антеридия переходит в аскогон, который делится после этого на ряд клеток, одна из которых содержит дикарион. Эта клетка или непосредственно трансформируется в сумку (у мучнисторосяных, образующих только одну сумку в клейстотеции), или из нее развиваются аскогенные гифы, а на них – сумки. Одновременно формируется перидий клейстотеция. Наружный и внутренний слои перидия различаются морфологически и функционально. Наружный слой его состоит из толстостенных гиф и является защитным, а внутренний – из тонкостенных, быстро лизирующихся гиф – выполняет питательную функцию. Из наружных слоев перидия развиваются придатки различного строения. Их форма, так же как и число сумок в клейстотеции – характерный признак родов мучнисторосяных.  Представители класса сордариомицеты. Порядок гипокреальные объединяет виды, которые образуют перитеции в хорошо развитых стромах, либо на мицелиальных сплетениях. Стромы обычно мясистые, яркоокрашенные, распростертые по субстрату, подушковидные или головчатые. Стромы развиваются на субстрате – обычно на пораженных органах растения-хозяина или из склероциев (виды спорыньи – Claviceps). Большинство представителей порядка – паразиты на цветковых растениях. Наиболее распространенный из них спорыньяпурпурная. Часто встречается на ржи, тимофеевке, пырее, костре, поражает также пшеницу, особенно твердую, ячмень. На пораженных спорыньей соцветиях образуются склероции, имеющие вид рожков черно-фиолетового цвета. Склероции зимуют в почве, куда они попадают при уборке урожая с культурных злаков или дикорастущих злаков. Весной склероции прорастают несколькими головчатыми стромами. Стромы обычно красноватого цвета. По периферии головок стромы выступают верхушки перитеций. После выбрасывания из перитециев аскоспоры разносятся ветром и попадают на растения. Аскоспоры спорыньи заражают злаки в период цветения. Через несколько дней после заражения на растениях развивается конидиальная стадия гриба. имеющая собственное название сфацелия (Sphacelia).В завязи образуется плотная масса мицелия, покрытая слоем конидиеносцев, продуцирующих огромное количество мелких конидий, погруженных в капли «медвяной росы». Медвяная роса – сладковатая жидкость, имеющая неприятный запах и содержащая большое количество сахаров. «Медвяная роса» играет существенную роль в распространении конидий гриба. Привлеченные ею насекомые переносят конидии на здоровые растения. Конидии могут распространяться и каплями дождя, при трении колосьев друг о друга под действием ветра, а после высыхания капель «медвяной росы» они могут переноситься ветром. В пораженных цветках ткани завязи полностью разрушаются и замещаются мицелием гриба. Ко времени окончания цветения злаков и созревания зерна мицелий спорыньи уплотняется и превращается в склероций. Склероции развиваются медленно. Сначала они желтовато-коричневые, затем приобретают серо-фиолетовую или черно-фиолетовую окраску. Полное их созревание приурочено ко времени спелости зерна.
Вопросы для самоконтроляНазовите совершенную стадию развития сумчатых грибов. Схематично объяснить сумкообразование аскомицетов Назвать типы плодовых тел аскомицетов Назвать морфологические типы сумок или асков Дать краткую характеристику фитопатогенным сумчатым грибам Литература Силантьева М.М., Сперанская Н.Ю., Соколова Л.В. Микология: лабораторные занятия, самостоятельная работа студентов: учебное пособие по дисциплине «Ботаника».- Барнаул: РИО Алтайского ГАУ, 2015. – 100 с. Инелова, З.А.. Биоразнообразие растительного мира.- Алматы, 2013 Нестерова, С.Г.. Лабораторный практикум по "систематике растений". - Алматы, 2011 Ботаника. Курс альгологии и микологии.- М., 2007 Лемеза, Н. А. Альгология и микология. Практикум : учеб. пособие / Н. А. Лемеза. – Минск: Выш.школа, 2008. – 319 с. Гарибова, Л. В. Основы микологии: морфология и систематика грибов и грибоподобных организмов / Л. В. Гарибова, С. Н. Лекомце-ва. – М. : Товарищество научных изданий КМК, 2005. – 220 с. 10 Outline of ascomycota – 2007 [Electronic resource] / Ed. H. T. Lumbsch, S. M. Huhndorf – Myconet, vol. 13. – Mode of access: http://www.fieldmuseum.org/myconet/outline.asp. – Date of access: 18.08.2008 |

