Клеточный иммунный ответ
 Скачать 0.74 Mb. Скачать 0.74 Mb.
|

|
Рис. 5. 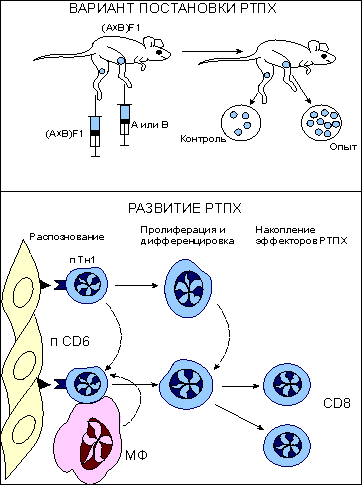 Реакция трансплантат против хозяина (РТПХ). Реакция трансплантат против хозяина (РТПХ). Верхняя часть рисунка демострирует вариант постановки РТПХ. Мышам- гибридам (А*В)F1 вводят лимфоциты одного из родителей (А или В) в подушечку одной из лап. Реципиент иммунологически толерантен к введенным клеткам, т.к. антигены родителей полностью представлены в гибриде. В подушечку противоположной лапы вводятся генетически идентичные клетки гибрида (контроль). Через 7 дней определяют массу или количество клеток в подколенном (региональном к месту введения) лимфатическом узле. Отношение количества клеток в "опытном" лимфатическом узле к количеству клеток в "контрольном" дает индекс РТПХ. При индексе более 1,3 реакция считается положительной. Нижняя часть рисунка указывает на клетки, принимающие участие в развитии реакции. Основными участниками являются предшественники цитотоксических Т-лимфоцитов (пCD8) и предшественники Т-клеток воспаления (пCD4). В процесс созревания включаются также макрофаги (МФ). Результатом реакции является накопление зрелых CD8. Увеличение количества клеток в лимфатическом узле или селезенке происходит не только за счет размножения введенных клеток, но и в результате привлечения в зону воспаления собственных клеток реципиента и их пролиферации in situ. Мышам (А*В)F1 вводят лимфоциты одного из родителей (А или В) в подушечку одной из лап. Реципиент иммунологически толерантен к введенным клеткам, так как антигены родителей полностью представлены в гибриде. Через 7 дней определяют массу или количество клеток в подколенном (регионарном к месту введения клеток) лимфатическом узле. Отношение числа клеток в "опытном" лимфатическом узле к числу клеток в "контрольном" узле дает индекс РТПХ. При отношении опыт:контроль, дающем индекс более 1,3, реакция считается положительной. Введенные чужеродные лимфоциты распознают неродственные антигены реципиента и формируют антигенспецифическую реакцию. В процесс распознавания включаются две субпопуляции лимфоцитов: предшественники CD8 T-клеток и предшественники CD4 T-клеток . Результатом реакции является накопление зрелых CD8 Т-клеток. Число клеток в селезенке или лимфатическом узле увеличивается не только за счет пролиферации введенных лимфоцитов, но и в результате привлечения в зону реакции собственных клеток реципиента. Реакция "трансплантат против хозяина" и поражение легких Одно из проявлений реакции "трансплантат против хозяина" - это прогрессирующие обструктивные нарушения вследствие поражения терминальных бронхиол, то есть облитерирующего бронхиолита . Он наблюдается у большинства реципиентов костного мозга и у 20-50% реципиентов легких и комплекса сердце-легкие. Бронхиолит протекает подобно острой инфекции, в рамках острой реакции отторжения, либо начинается незаметно и проявляется воспалением слизистых, постепенно нарастающим сухим кашлем , одышкой и обструктивными нарушениями . При рентгенографии грудной клетки обнаруживают усиление легочного рисунка и множественные мелкоочаговые тени , при гистологическом исследовании - очаги лимфоцитарной и нейтрофильной инфильтрации интерстициальной ткани и бронхиолит без васкулита . Лечение сводится к повышению доз иммунодепрессантов и назначению бронходилататоров и антибиотиков . Самое частое осложнение - вирусные инфекции , в частности цитомегаловирусная . Реакция "трансплантат против опухоли" Противоопухолевый эффект неочищенного аллотрансплантата, получивший название реакция "трансплантат против опухоли", обусловлен, по-видимому, Т-лимфоцитами и, возможно, NK-лимфоцитами донора. Во всяком случае, аллотрансплантат, очищенный от Т-лимфоцитов, таким эффектом не обладает. Имеется немало данных о том, что у реципиентов аллогенного костного мозга в искоренении злокачественной опухоли участвуют также иммунные механизмы. Конкретные медиаторы этой реакции, которая получила название " трансплантат против опухоли ", пока неизвестны. Реакции отторжения трансплантата Пересадка тканей или органов от одного индивидуума другому, генетически отличающемуся, или от одной инбредной линии мышей другой, также генетически отличной от линии донора, вызывает реакцию отторжения пересаженного биологического материала. Время отторжения первичного трансплантата - около 14 дней. Вторичный трансплантат отторгается быстрее: приблизительно за 5-7 дней. Отдельные реакции клеточного иммунитета, представленные выше, имеют свое интегральное проявление при отторжении чужеродной ткани. Собственно реакция отторжения включает два компонента: - специфический, связанный в основном с активностью цитотоксических T-клеток (CD8 T-клеток) , и - неспецифический, имеющий характер воспаления . Рис. 6. 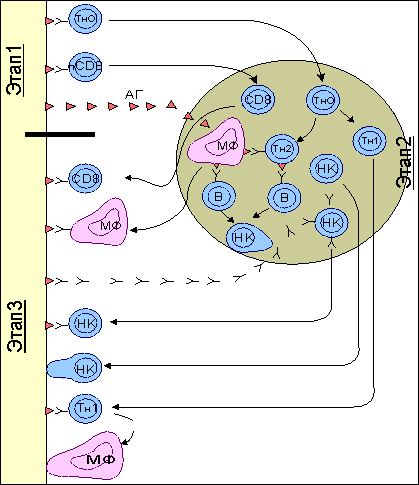 Реакция отторжения трансплантата. Реакция включает три этапа. На этапе I происходит распознавание антигенов трансплантата предшественниками цитотоксических Т-лимфоцитов (пCD8) и предшественниками хелперных и воспалительных Т-клеток (ТНО). После распознавания клетки мигрируют в ближайшую (региональную) лимфоидную ткань. В периферической лимфоидной ткани развиваются основные события, приводящие к формированию эффекторов реакции отторжения (этап II). пCD8 трансформируются в эффекторные зрелые цитотоксические Т-клетки (CD8). Свободные трансплантационные антигены, поступающие в лимфоидную ткань, захватываются антигенпрезентирующими клетками (отмечены только макрофаги - МФ) и подключают к ответу как ТН1-, так и ТН2-клетки. При совместном участии антигенпрезентирующих клеток, В-клеток и ТН2 формируется гуморальный иммунный ответ, являющийся дополнительным звеном отторжения. Здесь же происходит сорбция секретируемых антител на поверхности натуральных киллеров (НК), а также активация макрофагов либо под воздействием цитокинов Т-клеток, либо в результате сорбции антител. Активируются также и НК-клетки под воздействием цитокинов Т-лимфоцитов. На этапе III развиваются основные события трансплантационной реакции - отторжение чужеродной ткани. Отторжение реализуется при участии зрелых CD8 Т-клеток, активированных иммуноглобулинами макрофагов, антителами при участии комплемента, НК клетками, армированными иммуноглобулинами и активированными цитокинами При участии ТН1 в зону отторжения привлекаются макрофаги, обеспечивающие воспалительный компонент реакции отторжения. Развитие реакции трансплантационного иммунитета состоит из трех этапов ( рис. 6 ): - распознавание чужеродных антигенов трансплантата, - созревание и накопление эффекторов трансплантационной реакции отторжения в периферической, ближайшей к трансплантату лимфоидной ткани и - разрушение трансплантата. Этап I: распознавание. В процесс распознавания вступают предшественники цитотоксических Т-лимфоцитов и предшественники хелперных и воспалительных Т-клеток ( ТН0 ). После распознавания антигена клетки этих типов мигрируют в ближайшую лимфоидную ткань, например, в регионарный лимфатический узел . Этап 2: созревание и накопление. В периферической лимфоидной ткани развиваются основные события, приводящие к созреванию и накоплению клеток различных типов - эффекторов реакции отторжения. Предшественники цитотоксических Т-клеток , хелперных Т-клеток и Т-клеток воспаления дифференцируются в зрелые эффекторы. Процесс распознавания может происходить не только непосредственно в зоне трансплантата, но и в регионарной лимфоидной ткани за счет проникновения в нее антигенов трансплантата. В лимфоидной ткани антиген после усвоения макрофагами и выхода на клеточную поверхность в иммуногенной форме обеспечивает накопление T-клеток воспаления (ТН1) . Этот же антиген, экспрессируясь на поверхности В-клеток , включает хелперные Т-клетки (ТН2) , что обеспечивает накопление специфических антител . Таким образом, помимо эффекторов клеточного иммунитета в лимфоидной ткани идет процесс формирования эффекторов гуморального иммунного ответа. Секретируемые антитела могут сорбироваться на поверхности так называемых натуральных киллеров (НК-клеток) - особой популяции лимфоцитов, не имеющих маркеров Т- и В-клеток. Цитофильность антител к НК-клеткам обеспечивается взаимодействием Fc -участка иммуноглобулинов с соответствующим рецептором на поверхности НК-клеток. В результате НК-клетки, связавшие иммуноглобулин, приобретают способность к антителозависимому цитолизу клеток трансплантата. В процессе развития реакции на трансплантат присходит также активация макрофагов либо под воздействием цитокинов Т-клеток, либо в результате пассивной сорбции иммуноглобулинов на поверхности макрофагов по аналогии с НК-клетками. Этап III: разрушение. В разрушении и отторжении трансплантата участвуют перечисленные выше клеточные формы и специфические иммуноглобулины. Цитотоксические Т-лимфоциты и НК-клетки вступают в специфическую реакцию разрушения трансплантата: первые - за счет собственных антигенраспознающих рецепторов, вторые - за счет цитофильных антител. Клетки трансплантата неспецифически лизируются также НК-клетками, активированными цитокинами . Т-клетки воспаления после взаимодействия с антигенами трансплантата начинают активную секрецию хемотаксического макрофагингибирующего фактора , привлекающего в зону отторжения макрофаги, способные к неспецифическому лизису трансплантата (по своей форме это - типичная реакция воспаления ). Таким образом, в реакцию отторжения трансплантата включаются как специфические участники: CD8 Т-клетки, CD4 Т-клетки воспаления (ТН1), специфические иммуноглобулины, так и неспецифические: активированные макрофаги и натуральные киллеры. Генерация эффекторных Т-клеток. Общая характеристика. Первичное распознавание чужеродного антигена осуществляется функционально неподготовленными наивными Т-клетками , хотя внутритимусная дифференцировка уже определила основные свойства этих клеток, что отражается в их делении на субпопуляции. Созревание наивных Т-клеток в зрелые функционально активные (армированные) Т-клетки происходит в лимфоидной ткани . Место проникновения патогена (в более широком смысле - антигена) в организм, как правило, удалено от лимфоидных образований. С током лимфы антиген попадает в ближайшие лимфатические узлы . Если антиген оказывается в кровотоке, то он оседает в селезенке . Локализация антигена на слизистых приводит к его миграции в лимфоидную ткань, ассоциированную с этими слизистыми покровами: в пейеровы бляшки , миндалины . В периферической лимфоидной ткани имеются три типа специализированных клеток, которые способны усваивать антиген и представлять его в иммуногенной форме на своей поверхности для распознавания Т-клетками. Это - макрофаги , дендритные клетки и В-клетки ( табл. 1 и рис. 7 ). Все они получили общее название антигенпрезентирующих клеток (АПК) . Макрофаги не имеют гистологически определенного места локализации и широко представлены по всей лимфоидной ткани. Дендритные клетки связаны с Т-зоной лимфатических узлов. В-клетки концентрируются в фолликулах. Функция этих типов клеток - представление антигенных пептидов в комплексе с молекулами MHC класса I или II, т.е. придание проникшему антигену иммуногенных свойств. Рис. 7. 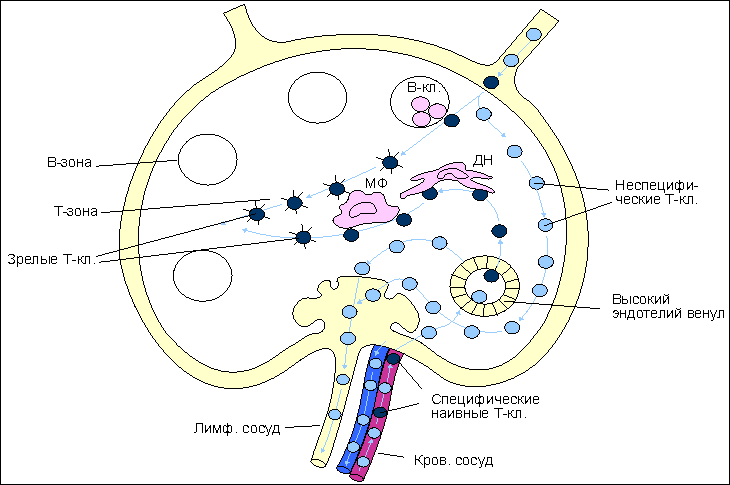 Рециркуляция Т-клеток после проникновения антигена в лимфоидную ткань. На рисунке представлена схема лимфатического узла с направлениями движения Т-клеток по органу. Антиген с током лимфы от места проникновения в организм заносится в лимфатический узел. Антигенпрезентирующие клетки (макрофаги - МФ, дендритные клетки - ДК, В-клетки - В-кл.) захватывают, перерабатывают и представляют антигенные пептиды в комплексе с молекулами I или II классов МНС на своей поверхности. Эти события активируют процесс рециркуляции клеток. Рециркулирующие наивные Т-клетки, среди которых имеются и специфичные к комплексу лимфоциты (черные кружки) попадают в лимфатический узел либо по афферентным лимфатическим сосудам, либо по кровеносным сосудам. В тех случаях, когда антиген поступает в орган с кровотоком, ему приходится преодолевать высокий эндотелий венул. Оказавшись в паренхиме органа, наивные антигенспецифические Т-клетки после распознавания имуногена на одной из антигенпрезентирующих клеток остаются в органе с тем, чтобы пройти дополнительную дифференцировку до функционально зрелых, армированных CD8 и CD4 Т-клеток. Антигеннеспецифические клетки, оказавшись невотребованными, покидают орган по эфферентному лимфатическому сосуду и вступают, таким образом, в новый цикл рециркуляции. Оказавшийся в лимфоидной ткани антиген провоцирует усиление рециркуляции лимфоцитов . Наивные Т-клетки попадают в лимфатические узлы в так называемую Т-зону через высокий эндотелий венул. Генерация зрелых (армированных) эффекторов Т-клеточного иммунного ответа начинается с распознавания антигенного пептида, комплексированного с молекулами I или II классов МНС, на поверхности макрофагов и дедритных клеток. Сам факт распознавания комплекса является обязательным, но недостаточным условием для инициации развития наивных Т-клеток в зрелые эффекторы. Необходимо предупреждающее включение кофакторов, которые способствуют взаимодействию антигенраспознающего рецептора Т-клеток с антигенным комплексом. Именно антигенпрезентирующие клетки обеспечивают такое двойное взаимодействие. Активация наивных Т-клеток при первичной встрече с антигеном получила название " примирование ". Это определение введено с тем, чтобы отличить первичное взаимодействие с антигеном от взаимодействия зрелых эффекторов с тем же антигеном, когда собственно и проявляется функциональное предназначение различных субпопуляций Т-клеток. В ряде случаев, в частности при формировании специфической цитотоксической реакции, презентирующая антиген клетка может выступать и как объект распознавания, и как объект цитолитического действия CD8 T-клеток после их созревания. Среди наивных Т-лимфоцитов, проникающих в лимфоидную ткань и временно локализующихся в Т-зонах, только один из 105 оказывается способным к специфическому взаимодействию. Остальные постепенно перемещаются в медуллярную зону и через эфферентный лимфатический сосуд покидают орган, чтобы вновь оказаться в кровотоке и быть занесенным в другие лимфоидные образования организма. Антигенпрезентирующие клетки в активации наивных T-клеток Хоминг наивных Т-клеток в лимфатические узлы осуществляется в результате взаимодействия L-селектинов Т-клеток с муцин-подобными адрессинами сосудов ( CD34 , GlyCAM-1 ). При хоминге лимфоцитов в лимфоидную ткань слизистых в процесс включаются L-селектины и MAdCAM-1 эндотелия слизистых покровов. При хоминге Т-клеток через эндотелий в паренхиму органа происходит за счет другой группы адгезивных молекул: интегрина LFA-1 и членов суперсемейства иммуноглобулинов ICAM-1 , ICAM-2 , ICAM-3 ( табл. 2 ). Табл. 2. Адгезивные молекулы, участвующие в заселении лимфоидной ткани наивными Т-клетками
Проникновение наивных T-лимфоцитов в кортикальную зону приводит к их столкновению с антигенпрезентирующими клетками . На начальном этапе процесса, как это ни странно, отсутствует специфический компонент взаимодействия: взаимодействия антигенраспознающего T-клеточного рецептора с иммуногеном на поверхности антигенпрезентирующих клеток . В межклеточный контакт вступают LFA-1 и CD2 T-клеток, а также ICAM-1 , ICAM-2 , ICAM-3 и LFA-3 (CD58) антигенпрезентирующих клеток. Дифференциальная роль каждого из этих адгезинов не установлена. Возможно, что их одновременное присутствие на клеточной поверхности обеспечивает определенный "запас прочности". Известно, что отсутствие синтеза LFA-1 у людей не нарушает функционирования T-системы. Выпадение звена взаимодействия пары LFA-1:ICAM компенсируется более активным включением пары CD2:LFA-3. Преходящее (временное) взаимодействие наивных Т-клеток с антигенпрезентирующими клетками осуществляется низкоаффинными LFA-1. Отсутствие жесткой формы контактных отношений между клетками имеет конкретный биологический смысл. Низкая аффинность адгезина позволяет Т-клетке приходить в контакт со многими антигенпрезентирующими клетками до тех пор, пока не произойдет специфическое узнавание иммуногена соответствующим антигенраспознающим рецептором T-клеток. Как только наивная Т-клетка находит "свой" иммуноген, ее перемещение в корковом слое приостанавливается. От Т-клеточного рецептора идет сигнал на LFA-1, меняющий конформацию адгезина, что приводит к усилению его аффинности по отношению к молекулам ICAM ( рис. 9.8 ). Механизм изменения аффинности LFA-1 неизвестен. Вероятно, в этом процессе принимают участие ионы Mg+2, но не Ca+2. Возникшие изменения стабилизируют контактные отношения между антигенспецифическими наивными Т-клетками и антигенпрезентирующими клетками. В результате такая Т-клетка становится подготовленной к пролиферации и дифференцировке в зрелые антигенспецифические Т-клетки. |