Клеточный иммунный ответ
 Скачать 0.74 Mb. Скачать 0.74 Mb.
|

|
ФГОУ ВПО Московская Государственная Академия Ветеринарной Медицины и Биотехнологии имени К. И. Скрябина Кафедра физиологии и этологии животных. Реферат на тему: «Клеточный иммунный ответ». Выполнила студентка 2 курса 4 группы Трефилова Е. А. Проверила Хомутинникова Юлия Александровна Москва 2006 Сожержание
Введение. Термин клеточный иммунитет (иммунитет, опосредованный клетками) используют для описания такого противоинфекционного или противоопухолевого иммунного ответа, в котором антителам (эффекторам гуморального иммунного ответа) принадлежит не ведущая, а вспомогательная роль. В отличие от В-системы иммунитета , которая нейтрализует антиген с помощью антител, Т-система иммунитета уничтожает антигены, представленные на клетках, через прямое взаимодействие субпопуляции T-клеток - специфических цитотоксических T-клеток (CD8 T-клеток, T-киллеров) с измененными собственными или чужеродными клетками. Вторая отличительная черта Т-клеток связана с особенностями распознавания антигена: Т-клетки распознают не собственно антигенный пептид ( эпитоп ), а его комплекс с молекулами I или II классов МНС . В тех случаях, когда антигенный пептид образует комплекс, включающий молекулы MHC класса I , распознавание и уничтожение осуществляется, как только что упоминалось выше, цитотоксическими CD8 T-клетками . В тех же случаях, когда антигенный пептид образует комплекс с молекулами MHC класса II , в процесс взаимодействия с таким комплексом вступают либо CD4 Т-клетки воспаления (ТН1) , либо хелперные CD4 Т-клетки (ТН2) . (Прежнее название клеток субпопуляции TH1 - Т-индукторы ). Если наивные Т-клетки распознают комплекс на поверхности макрофагов , поглотивших патоген, то такие клетки дифференцируются в CD4 Т-клетки воспаления (ТН1) , активируют эти макрофаги и тем самым способствуют внутриклеточному перевариванию (уничтожению) патогена. Путь проникновения антигена в клетку может осуществляться не только за счет неспецифической адгезии патогена на поверхности макрофагов, но и посредством специфического взаимодействия с предсуществующими антигенраспознающими рецепторами В-клеток (поверхностными иммуноглобулинами) . Экспрессия переработанного антигена в комплексе с молекулами II класса на поверхности В-клеток вызывает ответ наивных Т-клеток, дифференцирующихся в хелперные CD4 Т-клетки (ТН2) . В этом случае хелперные Т-клетки оказывают помощь В-клеткам в продукции антител , т.е. в формировании гуморального иммунного ответа . В табл. 1 представлены сравнительные характеристики особенностей проявления клеточного иммунного ответа и гуморального иммунного ответа. Табл. 1 Сравнительная характеристика проявлений клеточного и гуморального иммунного ответа
Основные реакции клеточного иммунитета. Реакции цитотоксических T-лимфоцитов При вирусной инфекции , опухолевом росте , отторжении трансплантата основными эффекторными клетками являются цитотоксические T-лимфоциты (CD8 T-клетки, T-киллеры) . Этапы цитолитического действия CD8 Т-клеток включают: - распознавание антигена предшественниками, - пролиферацию и дифференцировку T-клеток до зрелых эффекторов, - собственно процесс лизиса измененных своих вирусинфицированных и раковых клеток или чужеродных клеток трансплантированной ткани. Изучение клеточных и молекулярных механизмов, действующих на каждом этапе, было проведено, главным образом, в опытах in vitro. Приемы экспериментальной работы достаточно просты. Например, к суспензии клеток-мишеней, зараженных вирусом и меченных хромом-51, добавляют примированные к антигенам вируса лимфоциты, которые выделяют от мышей, предварительно иммунизированных данным вирусом. После определенного времени совместной инкубации (4-24 часа), в течение которого происходит лизис клеток-мишеней и выход в супернатант хрома-51, культуральные пробирки центрифугируют и определяют количество метки в супернатанте, являющееся эквивалентом числу разрушенных клеток-мишеней ( рис. 1 ). Рис. 1 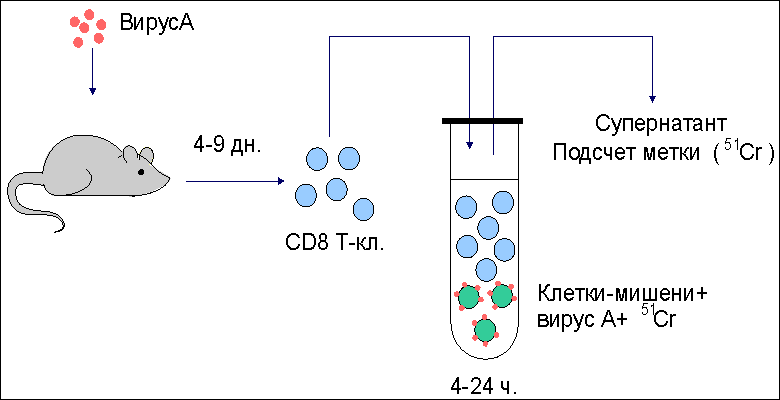 Схема одного из вариантов постановки цитотоксической реакции. От мышей, иммунизированных вирусом (условно, вирусом А), через 4-9 дней после введения антигена получают клетки лимфатического узла или селезенки. Оценку цитотоксической активности таких примированных клеток проводят in vitro c помощью клеток-мишеней, проинкубированных с вирусом и радиоактивной меткой 51Cr. К клеткам-мишеням добавляют в разном соотношении тестируемые клетки. В результате реакции цитолиза клеток- мишеней в супернатанте накапливается 51Cr. Количество метки в супернатанте является эквивалентом числу разрушенных клеток. Процесс лизиса чужеродных клеток состоит из нескольких этапов ( рис.2 ). Рис. 2. 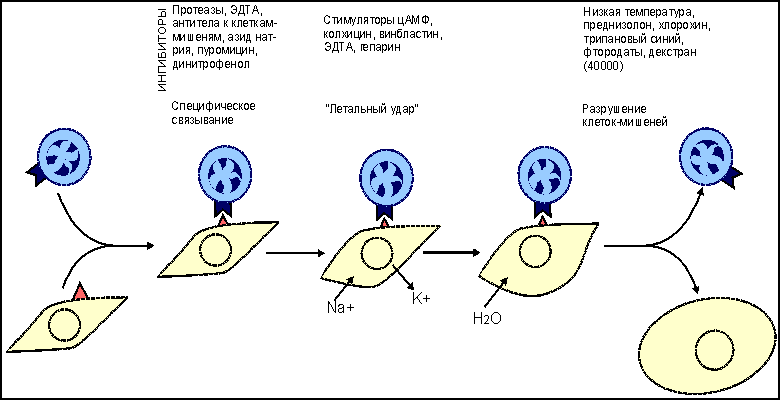 Этапы взаимодействия цитотоксических Т-лимфоцитов (ЦТЛ; Т-киллеров; CD8 Т-клеток) с клеткой-мишенью. Первый этап - специфическое связывание ЦТЛ с клеткой-мишенью. Механическое разобщение взаимодействующих клеток спасает клетку-мишень от лизиса. Второй этап - "летальный удар" - основное событие, предопределяющее гибель клетки-мишени. ЦТЛ должен находится в состоянии активного метаболизма. При этом жизнеспособность клетки-мишени не является обязательным условием. Механическое разобщение между взаимодействующими клетками на этом этапе не спасет клетку-мишень от гибели. Третий этап - заключительный, приводящий к лизису клетки-мишени. ЦТЛ остается неповрежденным и способен к дальнейшему цитотоксическому действию. Первый этап - специфическое связывание примированных CD8 T-клеток с поверхностным чужеродным антигеном (пептидами вирусных антигенов, трансплантационных антигенов, раковых антигенов). Взаимодействие антигенраспознающих рецепторов цитотоксических Т-клеток с чужеродным антигеном усиливается дополнительными неспецифическими молекулярными структурами клеточной поверхности, которые обеспечивают наиболее эффективную динамическую адгезию между клетками. Второй этап, получивший название "летального удара", представляет собой основное событие, предопределяющее гибель клетки-мишени. Механическое разобщение эффектора и клетки-мишени на этом этапе не спасает последнюю от гибели. Происходит характерное повышение проницаемости клеточной мембраны, нарушение баланса натрий-калиевого насоса . Механизм, лежащий в основе "летального удара", недостаточно ясен. Одним из факторов, повреждающих мембрану клетки, выступает лимфотоксин (бета-фактор некроза опухолей) . Третий этап, приводящий к лизису клетки-мишени, характеризуется увеличением ее объема за счет все большего проникновения молекул воды через поврежденную мембрану. В результате этих процессов происходит разрыв мембраны клетки-мишени и ее гибель. Эффекторная клетка сохраняется и способна к дальнейшему цитолитическому действию. Механизмы цитолитического действия цитотоксических Т-клеток Для изучения механизмов цитолитического действия цитотоксических T-лимфоцитов (CD8 T-клеток) используют два основных приема: - индукцию цитотоксических Т-лимфоцитов in vivo, когда для получения клеток-эффекторов иммунизируют животных антигенами клеток- мишеней (обычно это - аллогенные или опухолевые клетки); - индукцию таких клеток in vitro, когда созревание цитотоксических Т-клеток инициируют в смешанной культуре лимфоцитов (СКЛ). В последнем случае в качестве стимуляторов для примирования используют клетки лимфоидной ткани, генетически отличающиеся от распознающих предшественников CD8 Т-лимфоцитов модифицированные вирусом или иным антигеном (например, гаптеном ) сингенные клетки , а также сингенные раковые клетки. Клетки-стимуляторы облучают суперлетальной дозой или обрабатывают ингибитором клеточного деления для подавления их пролиферации. Предшественники CD8 Т-клеток вносятся в культуру интактными. В результате распознавания антигенов клеток-стимуляторов предшественники цитотоксических Т-лимфоцитов вступают в реакцию пролиферации и дифференцировки, что оценивается по включению 3Н-тимидина. Эффективность примирования предшественников CD8 Т-клеток оценивается во вторичной культуре по лизису клеток-мишеней, аутологичных или сингенных клеткам-стимуляторам ( рис. 3 ). Рис. 3. 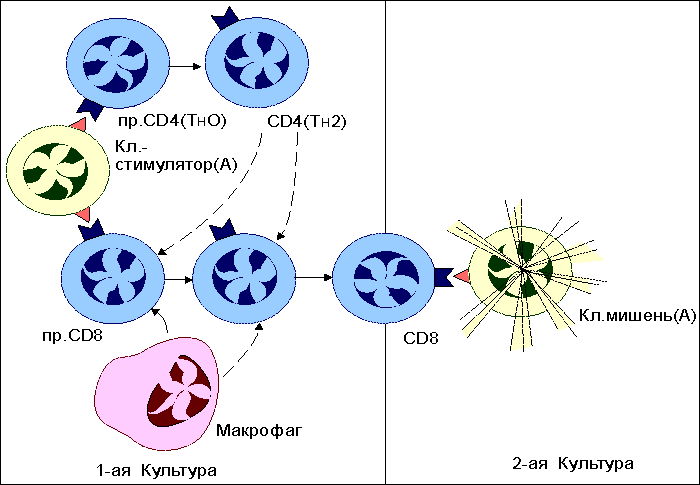 Реакция в смешанной культуре лимфоцитов (СКЛ). Эта реакция пролиферации развивается in vitro при взаимодействии генетически отличающихся (аллогенных) лимфоцитов. Результат реакции состоит в накоплении цитотоксических Т-лимфоцитов (CD8 Т-клеток), специфичных к антигенам гистосовместимости клеток-стимуляторов. В первичной культуре предшественники ЦТЛ (пр. CD8), представленные в суммарной популяции анализируемых лимфоцитов, после распознавания аллоантигенов клеток-стимуляторов вступают в процесс пролиферации и дифференцировки до зрелых ЦТЛ (CD8). Интенсивность пролиферативной реакции оценивают по включению 3Н-тимидина в размножающиеся клетки. Для созревания пр.CD8 необходима помощь со стороны макрофагов и хелперных CD4 Т-клеток (ТН2), образующихся из антигенраспознающих предшественников (ТН0). Оценку активности накопившихся в первичной культуре CD8 Т-клеток проводят во вторичной культуре (см. рис. 1 ). В качестве мишеней используют те аллогенные клетки, которые в первичной культуре выступали стимуляторами. Установлено, что распознавание антигена предшественниками CD8 Т-клеток недостаточно для генерации функционально активных цитотоксических лимфоцитов: необходима помощь со стороны хелперных CD4 T-клеток и макрофагов . Реакции гиперчувствительности замедленного типа Одной из форм клеточного иммуного ответа является реакция гиперчувствительности замедленного типа (ГЗТ) . Реакция впервые была описана немецким бактериологом Робертом Кохом в 1891 году. Введение интактным морским свинкам вирулентных бацилл туберкулеза (Mycobacterium tuberculosis) приводит к быстрому распространению возбудителя по организму, развитию острого инфекционного процесса , как правило, с летальным исходом. Картина меняется, если бациллы вводятся предварительно иммунизированным животным. В этом случае внутрикожно введенный возбудитель оказывается локализованным в месте инъекции в результате развития воспалительной реакции. Клетки воспалительного узелка представлены, в основном, макрофагами и лимфоцитами . Картину локальной воспалительной реакции можно наблюдать при введении не только бацилл, но и их продуктов - туберкулина и РРD (англ. "purified protein derivative"). Туберкулин - фильтрат культуральной жидкости M.tuberculosis. PPD состоит из белков, выделенных из культуры. Реакция локального воспаления развивается не только на бациллы туберкулеза, но и на широкий набор бактерий, вирусов, грибков, для которых макрофаги являются местом выживания и размножения. Кроме того, реакция может быть воспроизведена с модельными антигенами, например с эритроцитами различных видов животных. При повторном введении антигена реакция ГЗТ проявляется не сразу, а через 24-48 часов, отсюда и ее название - гиперчувствительность замедленного типа. По величине образующегося узелкового уплотнения (гранулемы) в месте введения антигена судят о напряженности иммунной реакции. Первичное введение антигена в интактный организм приводит к его захвату макрофагами, внутриклеточной деградации части антигена и экспрессии образующихся антигенных фрагментов (пептидов) на клеточной поверхности в комплексе с молекулами II класса МНС ( рис. 4 ). Рис. 4. 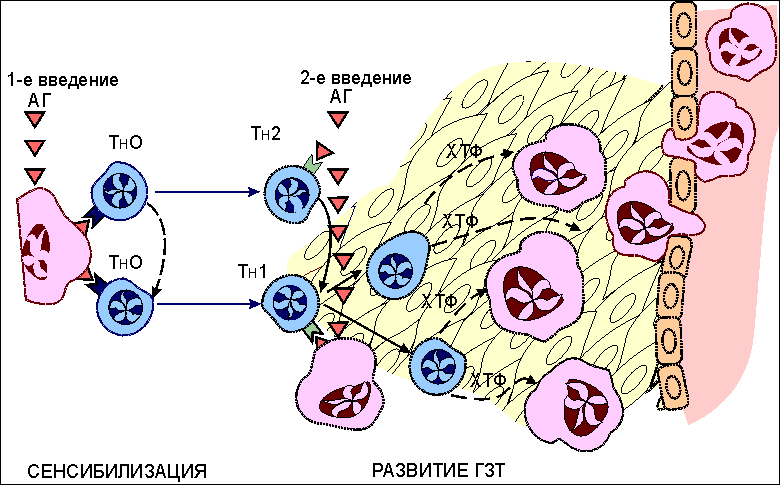 Реакция гиперчувствительности замедленного типа (ГЗТ). Гиперчувствительность замедленного типа - воспалительная реакция повышенной чувствительности, развивающаяся через 24-48 часов на месте повторного проникновения антигена. Первичное введение антигена в интактный организм приводит к его поглощению антигенпрезентирующими клетками (в частности, макрофагами), внутриклеточному разрушению и экспрессии образующихся антигенных фрагментов (пептидов) на клеточной поверхности в комплексе с молекулами II класса МНС. Иммуногенный комплекс распознается наивными CD4 Т-клетками (ТН0), которые дифференцируются в CD4 Т-клетки воспаления (ТН1; прежнее обозначение - TГЗТ, Т-индукторы). При повторной встрече с тем же антигеном зрелые ТН1 вступают в реакцию распознавания данного антигена. Результатом распознавания является активная продукция ТН1 хемотаксических факторов, привлекающих в зону проникновения антигена макрофаги и другие клетки воспаления из кровотока. Комплекс пептид:молекула II класса распознается наивными CD4 Т-клетками (ТН0) , которые дифференцируются в CD4 Т-клетки воспаления (ТН1) и хелперные CD4 Т-клетки (ТН2) . При повторном введении того же самого антигена дифференцированные ТН1 и ТН2 быстро вступают в реакцию взаимодействия с макрофагом , поглотившим антиген. Результатом специфического взаимодействия является секреция ТН1-клетками хемоаттрактантов (хемотаксических факторов) , привлекающих в зону проникновения антигена макрофаги из кровотока. Реакции трансплантат против хозяина (РТПХ) В клинической практике для компенсации врожденной иммунологической недостаточности или приобретенной недостаточности иногда вынуждены прибегать к пересадке клеток кроветворной и лимфоидной ткани. Поскольку в клеточном трансплантате содержатся иммунокомпетентные клетки, то, как правило, развивается реакция этих клеток на антигены реципиента. Реакция получила название реакции трансплантат против хозяина (РТПХ) . Для экспериментального воспроизведения РТПХ необходимо соблюдение следующих условий: 1) реципиент должен быть толерантным к введенным чужеродным клеткам; 2) трансплантируемые клетки должны обладать иммунологической компетенцией; 3) между клетками донора и реципиента должны существовать антигенные различия. В эксперименте реакцию оценивают либо по увеличению селезенки или лимфатических узлов , либо по смертности иммунологически инертного реципиента, которому введены лимфоциты генетически отличающегося донора. Один из вариантов РТПХ - увеличение массы и количества клеток в лимфатическом узле, регионарном к месту введения чужеродных лимфоцитов. Схема постановки реакции представлена на рис. 5 . | ||||||||||||||||||||||||