ЛЕКЦИИ Краткий курс лекций по медицинской микробиологии, вирусологии и иммунологии. Н.В. Рудаков 2002. ЛЕКЦИИ Краткий курс лекций по медицинской микробиологии, вирусол. Курс лекций по медицинской микробиологии, вирусологии и иммунологии. В 2 частях Учебное пособие. Омск, 2002. с
 Скачать 0.5 Mb. Скачать 0.5 Mb.
|

|
4.Морфологическая характеристика грибов. Грибы и простейшие имеют четко ограниченное ядро и относятся к эукариотам. Грибы крупнее бактерий, в эволюционном плане близки к растениям (наличие клеточной стенки, содержащей хитин или целлюлозу, вакуолей с клеточным соком, неспособность к перемещению, видимое движение цитоплазмы). Ядерный материал грибов отделен от цитоплазмы ядерной мембраной. Дрожжевые грибы образуют отдельные овальные клетки. Плесневые грибы формируют клеточные нитеподобные структуры- гифы. Мицелий- переплетение гифов- основная морфологическая структура. У низших грибов мицелий одноклеточный, не имеет внутренних перегородок (септ). Грибы размножаются половым и бесполым (вегетативным) способом. При вегетативном размножении образуются специализированные репродуктивные структуры- споры- конидии. Они могут располагаться в специализированных вместилищах- спорангиях (эндоспоры) или отшнуровываться от плодоносящих гиф (экзоспоры). Реже наблюдают образование спор внутри клеток (оидии), являющихся сегментами гиф. Дрожжевые клетки размножаются почкованием, мицелий не образуют. Половое размножение включает взаимодействие специализированных клеток, имеющих существенные различия в морфологии у различных грибов и часто используемых как дифференциально- диагностический признак. Для большинства видов грибов, имеющих медицинское значение, характерно наличие конидий (или экзоспор), являющихся формами неполового размножения. Их классификация во многом основывается на морфологических формах конидий. Их наиболее частые формы- бластопоры, хламидоспоры, артроспоры, конидиоспоры. Бластоспоры- простые структуры, котрые образуются в результате почкования, с последующим отделением почки от родительской клетки, например у дрожжевых грибов. Хламидоспоры образуются в результате увеличения гифальных клеток с образованием толстой оболочки, защищающей споры от неблагоприятных условий окружающей среды. Артроспоры- споры, образующиеся путем фрагментации гиф на отдельные клетки. Они встречаются у дрожжеподобных грибов, возбудителя кокцидиоидоза, тканевых форм дерматофитов в волосе, кожных чешуйках и в ногтях. Конидиоспоры- зрелые наружные споры, возникающие на дифференцированных конидиофорах (конидионосцах), отличающихся от других нитей мицелия по форме и размерам (у аспергилл, пеницилл) или располагающиеся по бокам и на концах любой ветви мицелия, прикрепляясь к ней непосредственно или тонкой ножкой. К эндоспорам совершенных грибов относятся спорангиоспоры мукоровых грибов, развивающихся в специальных органах (спорангиях), располагающихся на вершине спорангиеносца. Споры освобождаются при разрыве стенки спорангия. Эндоспоры обнаруживают также у тканевых форм возбудителей кокцидиоидоза. Они развиваются в круглых образованиях - сферулах, при разрыве стенки зрелой сферулы попадают во внешнюю среду. Основное функциональное отличие спор у бактерий и грибов: у бактерий споры обеспечивают переживание в неблагоприятных условиях окружающей среды, у грибов образование спор- способ размножения. 5. Морфологическая характеристика актиномицетов (лучистых грибов по старым классификациям). Актиномицеты - формы бактерий, имеющие истинный, не имеющий перегородок мицелий. Мицелиальный (в виде ветвящихся нитей) рост этих грамположительных бактерий придает им внешнее сходство с грибами. Это сходство усиливается вследствие наличия у высших форм актиномицетов наружных неполовых спор, которые называются конидиями. В отличие от грибов, актиномицеты имеют прокариотическое строение клетки, не содержат в клеточной стенке хитина или целлюлозы, размножаются только бесполым путем. У низших актиномицетов мицелий фрагментируется на типичные одноклеточные бактерии. Мицелий актиномицетов подразделяют на субстратный (в субстрате) и воздушный. К мицелиальным бактериям относят микобактерии, рода накардий и актиномицетов, несколько родов высших актиномицет. Представители рода Mycobacterium, в который входят возбудители туберкулеза, являются кислотоустойчивыми микроорганизмами, плохо воспринимающими краски. Их высокая резистентность во внешней среде , кислотоустойчивость и ряд других свойств связан с особым составом клеточной стенки, большим содержанием липидов и воска. У представителей родов Actinomyces и Nocardia мицелий выражен в значительно большей степени, чем у микобактерий, однако в старых культурах они также проявляют тенденцию фрагментироваться на отдельные клетки неправильной формы. Микроорганизмы рода Actinomyces являются анаэробами, Nocardia - аэробами, многие из которых проявляют кислотоустойчивость. Микроорганизмы, относящиеся к высшим актиномицетам (рода Streptomyces, Micromonospora) образуют мицелий и размножаются наружными неполовыми спорами или конидиями. Обычным местом обитания для большинства из них является почва. Однако ряд видов актиномицет и нокардий могут инфицировать раны и вызывать образование абсцессов. Для актиномицетов характерно образование друз - плотных “зерен” в гное, представляющих собой беспорядочно переплетенные в центре нити мицелия с радиально отходящими на периферию колбовидно расширенными на концах “дубинками”. С некоторыми актиномицетами (например, стрептомицетами) связана способность выработки антибиотиков. 6. Морфологическая характеристика простейших. Имеют эукариотическое строение клетки и значительно более сложную функциональную и морфологическую организацию по сравнению с бактериями и грибами. Снаружи тело простейших покрывает эластичная и ригидная пелликула, образованная внешним слоем цитоплазмы. У некоторых видов клеточная мембрана может включать опорные фибриллы и даже минеральный скелет. Простейшие могут иметь несколько ядер. Многие простейшие способны активно двигаться за счет псевдоподий, жгутиков или ресничек. Жизненный цикл паразитических простейших нередко включает образование промежуточных форм в различных хозяевах. Основные классы простейших: саркодовые или амебы- наиболее просто устроенные простейшие, споровики (малярийные плазмодии, токсоплазмы, пневмоцисты, бабезии), жгутиконосцы (трихомонады, лейшмании), инфузории. Простейшие очень широко распространены, достаточно сказать, что малярийными плазмодиями и токсоплазмами в сумме поражено до трети населения земного шара. Всего известно около 7 тысяч видов простейших, патогенных для различных растений, животных, человека, непатогенных- во много раз больше. Простейших изучает наука протозоология. Лекция № 3. Химическая структура, биохимические свойства и ферменты бактерий. Клетка- универсальная единица живой материи. По химическому составу существенных отличий прокариотических и эукариотических клеток нет. Химические элементы, входящие в состав живой материи, можно разделить на три основные группы. 1.Биогенные химические элементы (С, О, N, H). На их долю приходится 95% сухого остатка, в т.ч. 50%- C, 20%- O, 15%- N, 10%- H). 2.Макроэлементы- P, S,Cl, K, Mg, Ca, Na. На них приходится около 5 %. 3.Микроэлементы- Fe, Cu, I, Co, Mo и др. На них приходятся доли процента, однако они имеют важное значение в обменных процессах. Химические элементы входят в состав различных веществ- воды, белков, липидов, нейтральных жиров, углеводов, нуклеиновых кислот. Синтез соединений контролируется генами. Многие вещества бактериальная клетка может получать извне- из окружающей среды или организма хозяина. Вода составляет от 70 до 90 % биомассы. Содержание воды больше у капсульных бактерий, меньше всего- в спорах. Белки встречаются во всех структурных элементах клетки. Белки могут быть более простые (протеины) и сложные (протеиды), в чистом виде или в комплексе с липидами, сахарами. Выделяют структурные (структурообразующие) и функциональные (регуляторные) белки, к последним относятся ферменты. В состав белков входят как обычные для эукариотов аминокислоты, так и оригинальные- диаминопимелиновая, D-аланин, D-глютанин, входящие в состав пептидогликанов и капсул некоторых бактерий. Только в спорах находится дипиколиновая кислота, с которой связана высокая резистентность спор. Жгутики построены из белка флагеллина, обладающего сократительной способностью и выраженными антигенными свойствами. Пили (ворсинки) содержат особый белок- пилин. Пептидную природы имеют капсулы представителей рода Bacillus, возбудителя чумы, поверхностные антигены ряда бактерий, в том числе стафилококков и стрептококков. Белок А - специфический белок S.aureus - фактор, обусловлавливающий ряд свойств этого возбудителя. Белок М - специфический белок гемолитических стрептококков серогруппы А, позволяющий дифференцировать серовары (около 100), что имеет эпидемиологическое значение. Ряд белков содержит наружная мембрана грамотрицательных бактерий, из которых 3 - 4 мажорных (основных) и более 10- второстепенных, выполняющих различные функции. Среди мажорных белков- порины, образующие диффузные поры, через которые в клетку могут проникать мелкие гидрофильные молекулы. Белки входят в состав пептидогликана- биополимера, составляющего основу бактериальной клеточной стенки. Он состоит из остова (чередующиеся молекулы двух аминосахаров) и двух наборов пептидных цепочек- боковых и поперечных. Наличие двух типов связей- гликозидных (между аминосахарами) и пептидных, которые соединяют субъединицы пептидогликанов, придают этому гетерополимеру структуру молекулярной сети. Пептидогликан- наиболее устойчивое соединение, которое образует ригидную мешковидную макромолекулу, определяющую постоянную форму бактерий и ряд их свойств. 1.Пептидогликан содержит родо- и видоспецифические антигенные детерминанты. 2.Он запускает классический и альтернативный пути активации системы комплемента. 3.Пептидогликан тормозит фагоцитарную активность и миграцию макрофагов. 4.Он способен инициировать развитие гиперчувствительности замедленного типа (ГЗТ). 5.Пептидогликан обладает противоопухолевым действием. 6.Он оказывает пирогенное действие, т.е. вызывает лихорадку. Из соединений белков с небелковыми компонентами наибольшее значение имеют липопротеиды, гликопротеиды и нуклеопротеиды. Удивительное таинство жизни- синтез белка осуществляется в рибосомах. Существует два основных типа рибосом - 70S (S- константа седиментации, единица Сведберга) и 80S. Рибосомы первого типа встречаются только у прокариотов. Антибиотики не действуют на синтез белка в рибосомах типа 80S, распространенных у эукариотов. Липиды (главным образом форфолипиды) содержатся в цитоплазматической мембране (липидный бислой), в также в наружной мембране грамотрицательных бактерий. Есть микроорганизмы, содержащие большое количество липидов (до 40% сухого остатка)- микобактерии. В состав липидов входят различные жирные кислоты, весьма специфичные для разных групп микроорганизмов. Их определение имеет в ряде случаев диагностическое значение, например у анаэробов, микобактерий. У микобактерий туберкулеза в составе липидов имеется ряд кислотоустойчивых жирных кислот- фтионовая, миколовая и др. Высокое содержание липидов и их состав определяют многие свойства микобактерий туберкулеза: -устойчивость к кислотам, щелочам и спиртам; -трудная окрашиваемость красителями (используют специальные методы окраски, чаще- по Цилю- Нильсену); -устойчивость возбудителя к солнечной радиации и дезосредствам; - патогенность. Тейхоевые кислоты встречаются в клеточных стенках грамположительных бактерий. Представляют собой водорастворимые линейные полимеры, содержащие остатки глицерина или рибола, связанные фосфодиэфирными связыми. С тейхоевыми кислотами связаны главные поверхностные антигены ряда грамположительных бактерий. Углеводы встречаются чаще в виде полисахаридов, кторые могут быть экзо- и эндоклеточными. Среди экзоклеточных полисахаридов выделяют каркасные (входят в состав капсул) и истинно экзополисахариды (выходят во внешнюю среду). Среди бактериальных полисахаридов многие находят медицинское применение. Декстраны- полисахариды с большой молекулярной массой, по виду напоминают слизь. 6% раствор- кровезаменитель полиглюкин. Декстрановый гель сефадекс используется в колоночной хроматографии как молекулярное сито. Эндоклеточные полисахариды- запасные питательные вещества клетки (крахмал, гликоген и др.). Липополисахарид (ЛПС) - один из основных компонентов клеточной стенки грамотрицательных бактерий, это соединение липида с полисахаридом. ЛПС состоит из комплекса: 1.Липид А. 2.Одинаковое для всех грамотрицательных бактерий полисахаридное ядро. 3.Терминальная сахаридная цепочка (О- специфическая боковая цепь). Синонимы ЛПС- эндотоксин, О- антиген. ЛПС выполняет две основные функции- определяет антигенную специфичность и является одним из основных факторов патогенности. Это- эндотоксин, токсические свойства которого проявляются преимущественно при разрушении бактериальных клеток. Его токсичность определяется липидом А. ЛПС запускает синтез более 20 биологически активных веществ, определяющих патогенез эндотоксикоза, обладает пирогенным действием. Нуклеиновые кислоты- ДНК и РНК. Рибонуклеиновые кислоты (РНК) находятся главным образом в рибосомах (р-РНК- 80- 85%), т(транспортные)- РНК- 10%, м(матричные)- РНК- 1- 2%, главным образом в одноцепочечной форме. ДНК (дезоксирибонуклеиновая кислота) может находиться в ядерном аппарате (хромосомная ДНК) или в цитоплазме в специализированных образованиях- плазмидах- плазмидная (внехромосомная) ДНК. Микроорганизмы отличаются по структуре нуклеиновых кислот, содержанию азотистых оснований. Генетический код состоит всего из четырех букв (оснований) - А (аденин), Т (тимин), Г (гуанин) и Ц (цитозин). Наиболее часто для характеристики микроорганизмов используют как таксономический признак процентное соотношение Г/Ц, которое существенно отличается у различных групп микроорганизмов. Микроорганизмы синтезируют различные ферменты- специфические белковые катализаторы. У бактерий обнаружены ферменты 6 основных классов. 1.Оксидоредуктазы- катализируют окислительно- восстановительные реакции. 2.Трансферазы- осуществляют реакции переноса групп атомов. 3.Гидролазы- осущесвляют гидролитическое расщепление различных соединений. 4.Лиазы- катализируют реакции отщепления от субстрата химической группы негидролитическим путем с образованием двойной связи или присоединения химической группы к двойным связям. 5.Лигазы или синтетазы- обеспечивают соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле АТФ или аналогичного трифосфата. 6.Изомеразы - определяют пространственное расположение групп элементов. В соответствии с механизмами генетического контроля у бактерий выделяют три группы ферментов: - конститутивные, синтез которых происходит постоянно; - индуцибельные, синтез которых индуцируется наличием субстрата; - репрессибельные, синтез которых подавляется избытком продукта реакции. Ферменты бактерий делят на экзо- и эндоферменты. Экзоферменты выделяются во внешнюю среду, осуществляют процессы расщепления высокомолекулярных органических соединений. Способность к образованию экзоферментов во многом определяет инвазивность бактерий- способность проникать через слизистые, соединительнотканные и другие тканевые барьеры. Примеры: гиалуронидаза расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость тканей (клостридии, стрептококки, стафилококки и многие другие микроорганизмы); нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточном пространстве (холерный вибрион, дифтерийная палочка, вирус гриппа и многие другие). К этой же группе относятся энзимы, разлагающие антибиотики. В бактериологии для дифференциации микроорганизмов по биохимическим свойствам основное значение часто имеют конечные продукты и результаты действия ферментов. В соответствии с этим существует микробиологическая (рабочая) классификация ферментов. 1.Сахаролитические. 2.Протеолитические. 3.Аутолитические. 4.Окислительно- восстановительные. 5.Ферменты патогенности (вирулентности). Ферментный состав клетки определяется геномом и является достаточно постоянным признаком. Знание биохимических свойств микроорганизмов позволяет идентифицировать их по набору ферментов. Основные продукты ферментирования углеводов и белков- кислота, газ, индол, сероводород, хотя реальный спектр для различных микроорганизмов намного более обширный. Основные ферменты вирулентности- гиалуронидаза, плазмокоагулаза, лецитиназа, нейраминидаза, ДНК-аза. Определение ферментов патогенности имеет значение при идентификации ряда микроорганизмов и выявления их роли в патологии. Ряд ферментов микроорганизмов широко используется в медицине и биологии для получения различных веществ (аутолитические, протеолитические), в генной инженерии (рестриктазы, лигазы). Лекция № 4. Физиология и принципы культивирования микроорганизмов. Метаболизм микроорганизмов. Для роста и размножения микроорганизмы нуждаются в веществах, используемых для построения структурных компонентов клетки и получения энергии. Метаболизм (т.е. обмен веществ и энергии) имеет две составляющих- анаболизм и катаболизм. Анаболизм- синтез компонентов клетки (конструктивный обмен). Катаболизм- энергетический обмен, связан с окислительно- восстановительными реакциями, расщеплением глюкозы и других органических соединений, синтезом АТФ. Питательные вещества могут поступать в клетку в растворимом виде (это характерно для прокариот)- осмотрофы, или в виде отдельных частиц- фаготрофы. Основным регулятором поступления веществ в бактериальную клетку является цитоплазматическая мембрана. Существует четыре основных механизма поступления веществ: -пассивная диффузия- по градиенту концентрации, энергонезатратная, не имеющая субстратной специфичности; - облегченная диффузия- по градиенту концентрации, субстратспецифичная, энергонезатратная, осуществляется при участии специализированных белков пермеаз; - активный транспорт- против градиента концентрации, субстратспецифичен (специальные связывающие белки в комплексе с пермеазами), энергозатратный (за счет АТФ), вещества поступают в клетку в химически неизмененном виде; - транслокация (перенос групп)- против градиента концентрации, с помощью фосфотрансферазной системы, энергозатратна, вещества (преимущественно сахара) поступают в клетку в форфорилированном виде. Основные химические элементы- органогены, необходимые для синтеза органичеких соединений- углерод, азот, водород, кислород. В зависимости от источника потребляемого углерода микробы подразделяют на аутотрофы (используют CO2) и гетеротрофы (используют готовые органические соединения). В зависимости от источника энергии микроорганизмы делят на фототрофы (энергию получают за счет фотосинтеза- например, цианобактерии) и хемотрофы (энергия добывается за счет химических, окислительно- восстановительных реакций). Если при этом донорами электронов являются неорганические соединения, то это литотрофы, если органические- органотрофы. Если бактериальная клетка в состоянии синтезировать все необходимые для жизнедеятельности вещества, то это прототрофы. Если бактерии нуждаются в дополнительных веществах (факторах роста), то это ауксотрофы. Основными факторами роста для труднокультивируемых бактерий являются пуриновые и пиримидиновые основания , витамины, некоторые (обычно незаменимые) аминокислоты, кровяные факторы (гемин) и др. Дыхание микроорганизмов. Путем дыхания микроорганизмы добывают энергию. Дыхание- биологический процесс переноса электронов через дыхательную цепь от доноров к акцепторам с образованием АТФ. В зависимости от того, что является конечным акцептором электронов, выделяют аэробное и анаэробное дыхание. При аэробном дыхании конечным акцептором электронов является молекулярный кислород (О2), при анаэробном- связанный кислород ( -NO3 , =SO4, =SO3). Примеры. 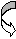 О2 О2А  эробное дыхание донор водорода H2O эробное дыхание донор водорода H2OАнаэробное дыхание н 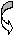 итратное окисление NO3 итратное окисление NO3(  факультативные анаэробы) донор водорода N2 факультативные анаэробы) донор водорода N2с 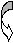 ульфатное окисление SO4 ульфатное окисление SO4(  облигатные анаэробы) донор водорода H2S облигатные анаэробы) донор водорода H2SПо типу дыхания выделяют четыре группы микроорганизмов. 1.Облигатные (строгие) аэробы. Им необходим молекулярный (атмосферный) кислород для дыхания. 2.Микроаэрофилы нуждаются в уменьшенной концентрации (низком парциальном давлении) свободного кислорода. Для создания этих условий в газовую смесь для культивирования обычно добавляют CO2, например до 10- процентной концентрации. 3.Факультативные анаэробы могут потреблять глюкозу и размножаться в аэробных и анаэробных условиях. Среди них имеются микроорганизмы, толерантные к относительно высоким (близких к атмосферным) концентрациям молекулярного кислорода - т.е. аэротолерантные, а также микроорганизмы которые способны в определенных условиях переключаться с анаэробного на аэробное дыхание. 4.Строгие анаэробы размножаются только в анаэробных условиях т.е. при очень низких концентрациях молекулярного кислорода, который в больших концентрациях для них губителен. Биохимически анаэробное дыхание протекает по типу бродильных процессов, молекулярный кислород при этом не используется. Аэробное дыхание энергетически более эффективно (синтезируется большее количество АТФ). В процессе аэробного дыхания образуются токсические продукты окисления (H2O2- перекись водорода, -О2 - свободные кислородные радикалы), от которых защищают специфические ферменты, прежде всего каталаза, пероксидаза, пероксиддисмутаза. У анаэробов эти ферменты отсутствуют, также как и система регуляции окислительно- восстановительного потенциала (rH2). Основные методы создания анаэробных условий для культивирования микроорганизмов. 1.Физический- откачивание воздуха, введение специальной газовой безкислородной смеси (чаще- N2- 85%, CO2- 10%, H2- 5%). 2.Химический- применяют химические поглотители кислорода. 3.Биологический- совместное культивирование строгих аэробов и анаэробов (аэробы поглощают кислород и создают условия для размножения анаэробов). 4.Смешанный- используют несколько разных подходов. Необходимо отметить, что создание оптимальных условий для строгих анаэробов- очень сложная задача. Очень непросто обеспечить постоянное поддержание безкислородных условий культивирования, необходимы специальные среды без содержания растворенного кислорода, поддержание необходимого окислительно- восстановительного потенциала питательных сред, взятие и доставка, посев материала в анаэробных условиях. Существует ряд приемов, обеспечивающих более подходящие условия для анаэробов- предварительное кипячение питательных сред, посев в глубокий столбик агара, заливка сред вазелиновым маслом для сокращения доступа кислорода, использование герметически закрывающихся флаконов и пробирок, шприцев и лабораторной посуды с инертным газом, использование плотно закрывающихся эксикаторов с горящей свечой. Используются специальные приборы для создания анаэробных условий- анаэростаты. Однако в настоящее время наиболее простым и эффективным оборудованием для создания анаэробных и микроаэрофильных условий является система “Газпак” со специальными газорегенерирующими пакетами, действующими по принципу вытеснения атмосферного воздуха газовыми смесями в герметически закрытых емкостях. Основные принципы культивирования микроорганизмов на питательных средах. 1.Использование всех необходимых для соответствующих микробов питательных компонентов. 2.Оптимальные температура, рН, rH2, концентрация ионов, степень насыщения кислородом, газовый состав и давление. Микроорганизмы культивируют на питательных средах при оптимальной температуре в термостатах, обеспечивающих условия инкубации. По температурному оптимуму роста выделяют три основные группы микроорганизмов. 1.Психрофилы- растут при температурах ниже +20 градусов Цельсия. 2.Мезофилы- растут в диапозоне температур от 20 до 45 градусов (часто оптимум- при 37 градусах С). 3.Термофилы- растут при температурах выше плюс 45 градусов. Краткая характеристика питательных сред. По консистенции выделяют жидкие, плотные (1,5- 3% агара) и полужидкие (0,3- 0,7 % агара) среды. Агар- полисахарид сложного состава из морских водорослей, основной отвердитель для плотных (твердых) сред. В качестве универсального источника углерода и азота применяют пептоны- продукты ферментации белков пепсином, различные гидролизаты- мясной, рыбный, казеиновый, дрожжевой и др. По назначению среды разделяют на ряд групп: - универсальные (простые), пригодные для различных нетребовательных микроорганизмов (мясо- пептонный бульон- МПБ, мясо- пептонный агар- МПА); - специальные- среды для микроорганизмов, не растущих на универсальных средах (среда Мак- Коя на туляремию, среда Левенштейна- Иенсена для возбудителя туберкулеза); - дифференциально- диагностические- для дифференциации микроорганизмов по ферментативной активности и культуральным свойствам ( среды Эндо, Плоскирева, Левина, Гисса); - селективные (элективные)- для выделения определенных видов микроорганизмов и подавления роста сопутствующих- пептонная вода, селенитовая среда, среда Мюллера. По происхождению среды делят на естественные, полусинтетические и синтетические. Рост и размножение микроорганизмов. Бактериальные клетки размножаются в результате деления. Основные стадии размножения микробов в жидкой среде в стационарных условиях: - лаг- фаза (начальная стадия адаптации с медленным темпом прирости биомассы бактерий); - экспоненциальная (геометрического роста) фаза с резким ростом численности популяции микроорганизмов (2 в степеии n); - стационарная фаза (фаза равновесия размножения и гибели микробных клеток); - стадия гибели - уменьшение численности популяции в связи с уменьшением и отсутствием условий для размножения микроорганизмов (дефицит питательных веществ, изменение рH, rH2, концентрации ионов и других условий культивирования). Данная динамика характерна для периодических культур с постепенным истощением запаса питательных веществ и накоплением метаболитов. Если в питательной среде создают условия для поддержания микробной популяции в экспоненциальной фазе- это хемостатные (непрерывные) культуры. Характер роста бактерий на плотных и жидких питательных средах: сплошной рост, образование колоний, осадок, пленка, помутнение. Чистая культура- популяция одного вида микроорганизмов. Основные принципы получения чистых культур: механическое разобщение, рассев, серийные разведения, использование элективных сред, особых условий культивирования (с учетом устойчивости некоторых микробов к определенным температурам, кислотам, щелочам, парциальному давлению кислорода, рН и мн.др). |