Фитоценоз. Курсовая работа научный регистрации по журналу учета курсовых работ
 Скачать 350.6 Kb. Скачать 350.6 Kb.
|

|
МИНОБРНАУКИ РОССИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ «БАШКИРСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМ.М.АКМУЛЛЫ» ЕСТЕСТВЕННО–ГЕОГРАФИЧЕСКИЙ ФАКУЛЬТЕТ Кафедра биоэкологии и биологического образования Направление Биология Курс II ВРЕМЕННАЯ СТРУКТУРА РАСТИТЕЛЬНЫХ СООБЩЕСТВ КУРСОВАЯ РАБОТА Научный руководитель: № регистрации по журналу учета курсовых работ ___________ Дата защиты___________________ Оценка________________________ подпись руководителя Уфа 2018 Содержание
ВВЕДЕНИЕ Актуальность исследования. Изучение растительных сообществ является важной составляющей в понимании процессов, происходящих в биогеоценозах (Разумовский, 1981). Неоднородность и динамичность среды в большинстве растительных сообществ создает условия, при которых ни один вид растений не может полностью использовать наличные ресурсы энергии и вещества и тем самым исключить возможность участия в сообществах других видов растений. Как следствие этого в течение очень длительного периода времени шел отбор видов растений и животных, включая микроскопические организмы, способные существовать совместно в определенных условиях среды. Создавались биогеоценозы, в которых каждый вид занимал определенное место – экологическую нишу. При этом количественные соотношения видов изменялись в определенных пределах в соответствии с изменением условий их обитания в течение года и год от года. Формировались растительные сообщества из растений, использующих различные горизонты среды (надземной и подземной), с разной интенсивностью в отдельные годы и периоды года (Василевич, 1983). Под влиянием физико–географических условий (климат, почва, влажность и пр.) и борьбы за существование растительные виды образуют естественные группировки или растительные сообщества (Быков, 1983). Для каждого растительного сообщества (фитоценоза), можно определить состав входящих в него видов – флористический состав. В таких сообществах разные виды представлены разным количеством экземпляров, который является относительно постоянным (Прокопьева, 2009). Целью работы: изучение структуры растительного мира. Задачи исследования: Изучить организацию растительных сообществ. Рассмотреть понятие фитоценоз, а также его классификацию и динамику. Структура работы: Курсовая работа изложена на 26 страницах, состоит из введения, 2 глав, заключения, списка использованной литературы. Работа проиллюстрирована 7 рисунками, список использованных источников включает 9 наименований. ГЛАВА 1. ОРГАНИЗАЦИЯ РАСТИТЕЛЬНЫХ СООБЩЕСТВ (ФИТОЦЕНОЗОВ) 1.1. Флористический состав При описании растительных сообществ прежде всего выявляется флористический состав. Видовое богатство растительного покрова определяется причинами как глобального, так и локального масштаба. Зависимость видового богатства от географической широты показана в работе В.М. Шмидта (1981). От полуострова Канин (68° с.ш., 45° в. д.) до Латвии (58° с.ш., 25° в. д.) число видов конкретных флор увеличивается в 3 раза. Этот рост обусловлен не только широтой, но и долготой местности, т. е. уменьшением континентальности. Другой экотопический фактор, а именно эдафический, также оказывает существенное влияние. В качестве примера можно привести поемные луга. Как известно, условия поймы весьма благоприятны для развития луговой растительности, но такие мощные факторы, как длительность затопления при паводках и аллювиальность, ведут довольно жесткий отбор видов. Так, в прирусловой зоне поймы реки Луги (Ленинградская область), где наилок в отдельные годы может достигать толщины 15 см, мы находим в основном виды растений, способные противостоять ежегодным наносам. Это длиннокорневищные растения, способные быстро прорастать через наносы (Bromopsis inermis, Phalaroides arundinacea и др.). Число видов здесь не превышает 10 – 12, в то время как в средней части поймы в сходных условиях увлажнения, но при меньшем аллювии можно встретить 35–40 видов. Таким образом, при довольно богатом видовом составе пойменных лугов сообщества прирусловой части бедны во флористическом отношении (Прокопьева, 2009). Важная роль в регулировании видового состава принадлежит биотопу, особенно если сообщество образовано сильным эдификатором. Ярким примером этому может служить еловый лес. Под пологом елового леса формируются условия, ограничивающие видовое разнообразие, хотя пространство там занято не полностью. Так, в ельнике кислично–зеленомошном на площади 100 м2 можно встретить не более 10–15 видов, включая мхи; еще меньше видов в чернично–сфагновых ельниках. В данном случае именно биотоп приводит к обеднению флористического состава (экотоп позволяет существовать здесь большему числу видов). В других случаях, напротив, биотопические особенности способствуют обогащению видового состава. Примером могут быть лиственные леса, где световой режим позволяет весной развиваться синузии эфемероидов, отсутствующей в хвойных лесах. Вообще переменность экологического режима, какими бы причинами она ни вызывалась, ведет к увеличению видового разнообразия (Шенников, 1964). Существенное влияние на количество видов в сообществах оказывают человек и животные. Так, в степных сообществах флора обогащается в результате роющей деятельности грызунов: на выбросах поселяются корневищные виды (Agropyron pecitnatum. Leymus ramosus), а также эфемеры и сорняки (Ranunculus orthoceras, Lepidtum perfoliatum и др.), плохо переносящие задернение в целинной степи. Выпас скота на лугах также изменяет видовой состав: выпадают виды, не переносящие вытаптывания, и разрастаются пастбищники – лапчатка (Poientilla ansenna), манжетка (Alchemilla sp.) и др. Многократное выкашивание луга ранее срока созревания семян приводит к обеднению его флоры, особенно разнотравья. Таковы основные факторы, определяющие видовое разнообразие растительных сообществ (Василевич, 1983). В зависимости от условий сообщества могут быть флористически простые и сложные, флористически богатые и бедные. Простые бедные сообщества – почти чистые заросли прибрежноводных растений: тростника (Phragmites australis), камыша (Sctrpus lacustns) и др. В видовом отношении примером богатых сообществ служит влажный тропический лес. Богаты видами северные или луговые степи. Так, видовая насыщенностъ луговых степей достигает 50 и более видов сосудистых растений на 1м2 . Здесь большое количество злаков (Stipa joannis, St.stenophylla, Poa angustifolia, Festuca sulcata и др.), обилие бобовых и особенно разнотравья (виды p.Trifolium, Salvia pratensis, Fihpendula hexapetala, виды p.Galium) (Разумовский, 1981). Встает вопрос, почему формируются многовидовые сообщества, несмотря на то, что их существование как будто противоречит представлениям о конкурентных взаимоотношениях. Есть мнение, что эти сообщества более устойчивы и более полно используют природные ресурсы, что и обеспечило их формирование в процессе эволюции. Справедливо подвергая критике эти соображения, В. И. Василевич (1979) приходит к выводу, что процесс видообразования идет независимо от ценотических процессов и отнюдь не направлен на создание рационально устроенных растительных сообществ. Возникают виды – экологические дубли, с точки зрения фитоценологии – фитоценотически избыточные виды. Процессы же конкуренции, по мнению В. И. Василевича, протекают медленно и в ограниченной зоне, что и дает возможность сосуществовать нескольким видам в одной экологической нише. Относительно видового богатства общим положением можно считать следующее: чем ближе условия к экстремальным, тем беднее во флористическом отношении растительный покров. Это – уже упоминавшиеся заросли прибрежено–водных растений, сообщества солянок на солончаках, растительность такыров и движущихся песков. Выявление флористического состава, как уже говорилось, осуществляется на начальном этапе любого геоботанического исследования. Однако флористический список недостаточен для характеристики сообщества (Уиттекер, 1980). 1.2. Основные свойства фитоценозов Фитоценоз как система представляет сложное образование, которое следует рассматривать как своеобразную природную систему. Любой фитоценоз состоит из различных компонентов, каким-то образом связанных друг с другом. Эти связи между различными видовыми группами бывают различные и поэтому имеют определенную устойчивость системы. При очень слабых связях, система может быть разрушена и такие фитоценозы нельзя назвать устойчивыми, они могут быть обречены на смену (Разумовский, 1981). Структура и состав фитоценозов зависит от взаимоотношений растений между собой и с окружающей средой. Под средой обитания понимают комплекс абиотических, биотических, антропогенных факторов. Абиотические факторы (неживой природы) подразделяют на климатические, орографические, гидрологические, биотические – на фитогенные (влияние растений) и зоогенные (влияние животных) и почвенные. Антропогенные факторы – прямые и косвенные воздействия, оказываемые человеком в результате своей деятельности. Абиотические, биотические и антропогенные факторы, воздействующие на организм в процессе его онтогенеза, называют экологическими факторами, а науку, изучающую взаимоотношения организма со средой – экологией (Шенников, 1964). Кроме понятия «среда обитания», есть понятие «условия существования», означающее совокупность жизненно необходимых факторов среды (свет, тепло, влага, питательные вещества). В геоботанике часто используют термины «местообитание» и «местонахождение», введенные еще А. Декандолем. Под местообитанием он понимал совокупность природных условий данной местности, где произрастает изучаемая особь или вид растений. Местонахождение – географический пункт, местность, где найдена особь или вид (Прокопьева, 2009). Экотоп – определенная территория или акватория со свойственными особенностями почв, грунтов, микроклимата и других факторов в неизмененном организмами виде. В результате воздействия на экотоп растений, животных, микроорганизмов экотоп превращается в биотоп. Каждому биоценозу свойственен свой биотоп. При одном экотопе в разных биоценозах формируются разные биотопы (Прокопьева, 2009). Растения представляют требования к среде обитания в соотвествии со своей наследственностью, обусловленной их происхождением из определенных природных зон. 1.3. Классификация фитоценозов Растительный покров разнообразен, и поэтому, чтобы правильно учитывать и использовать растительные ресурсы, необходимо все это многообразие привести в определенную систему, т.е. классифицировать. Следует различать классификацию флоры и растительного покрова. Основы классификации флоры заложены шведским ученым Карлом Линнеем. Он описал около 1200 родов растений, установил более 8000 видов. Для видов растений К. Линней предложил двойные названия (бинарную номенклатуру) на латинском языке. В названии вида первое слово (существительное) обозначает род, второе (прилагательное) обозначает вид. Например, клевер луговой (TrifoliumpratanseL.). Буква «L» означает, что вид описан К. Линнеем. Двойные названия указывают на родство между видами, происхождение их от общего предка. Сходные роды объединяют в семейства. Так, род клевер вместе с родами люцерна (Medicago), чина (Lathurus), горох (Pisum) и другими относится к семейству бобовых (Zeguminosae). Семейства объединяются в порядки, порядки — в классы, классы — в отделы (типы) (Александрова, 1969). Систематика растений, созданная К. Линнеем, была в значительной мере искусственной, так как он недостаточно учел родство видов, что объясняется малой изученностью растительности, в то время. Впоследствии учеными разных стран была создана естественная система растений. В настоящее время для установления родства между видами учитывают не только весь комплекс эколого-морфологических признаков растений, но также изучают их генотип, в частности набор хромосом. Широко используют достижения биохимии (хемосистематика). Родственные виды, роды и семейства имеют сходный химический состав. Это учитывается, в частности, при поиске лекарственных растений (Василевич, 1983). Основная единица классификации растительного покрова — ассоциация. В 1910 г. на Брюссельском ботаническом конгрессе было принято следующее определение ассоциации: «Ассоциация есть растительное сообщество определенного флористического состава с особыми условиями существования, особой физиономией», т. е. ассоциация — это тип фитоценоза. Фитоценоз — понятие конкретное, он приурочен к определенной территории. Ассоциация же как тип фитоценоза абстрактна. Например, ассоциация «ельник кисличный» характерна для многих районов южной тайги и хвойно-широколиственных лесов. Все ельники-кисличники сходны по господствующим в их ярусах и синузиях видам растений. Ярусы древостоя образованы елью, подлесок состоит из рябины, крушины, ив и др.; в травяно-кустарничковом покрове господствует кислица, омоховение слабое, преобладает мох плеурозий Шребера (Быков, 1983). Классификационные системы растительности в России строятся по принципу фитоценотического сходства, выражающегося в наличии в сообществах общих доминантов, эдификаторов и жизненных форм. При выделении единиц более высокого ранга учитывается экологическая и физиологическая близость эдификаторов (Александрова, 1969). В России принято выделять следующие таксономические категории растительности: ассоциация, группа ассоциаций, формация, группа формаций, класс формаций, тип растительности. Иногда применяют также таксон — класс ассоциаций и некоторые другие. К ассоциации относят фитоценозы с однородным видовым составом, одинаковой структурой, приуроченностью к сходным условиям местообитания. Ассоциации выделяют по однородности видового состава, но не по полной общности. Общими должны быть доминанты и содоминанты (Дохман, 1973). К группе ассоциаций относят все ассоциации, различающиеся по составу одного из ярусов при тождестве основных особенностей остальных ярусов, в том числе главного яруса: Так, группа ассоциаций ельники зеленомошные объединяет ассоциации ельников, в мохово-лишайниковом покрове которых преобладают зеленые мхи. В ее состав входят ассоциации ельник бруснично-зеленомошный, чернично-зеленомошный, кисличный зеленомошный и зеленомошный чистый (без кустарничков). В формацию входят группы ассоциаций, характеризующиеся общими эдификаторами. Так, в таежных лесах различают формации: ели европейской и сибирской, сосны обыкновенной, березы повислой. Формация — основная таксономическая единица среднего ранга. К группе формаций относят такие формации, эдификаторы которых принадлежат к одной жизненной форме. Так, формации ели сибирской и европейской, пихты сибирской и других теневыносливых хвойных деревьев образуют группу формаций темнохвойные леса. А формации светолюбивых хвойных деревьев (сосны обыкновенной, лиственницы сибирской др.) составляют группу формаций светлохвойные леса. Группы формаций с эдификаторами, сходными по жизненной форме, объединяют в классы формаций. Так, группы формаций темнохвойных и светлохвойных лесов объединяют в класс формаций хвойные леса. Группы формаций мелколиственных и широколиственных лесов умеренной зоны образуют класс формаций лиственные леса с опадающей на зиму листвой. Классы формаций объединяют в тип растительности. Наиболее правильно тип растительности выделять по морфолого-экологическим признакам. Классы формаций хвойные и лиственные леса с опадающей на зиму листвой относят к лесному типу растительности (леса). Выделяют следующие основные типы растительности: лесная, болотная, луговая, степная, пойменная. Рассмотренная выше система классификации растительности— субординационная, так как представлена рядом таксонов, последовательно подчиняющихся один другому. Название ассоциации дают на русском и латинском языках по доминантным видам ярусов растительности в сообществе. Например, если в древостое леса господствует сосна, в травяно-кустарничковом покрове — вереск, а в мохово-лишайниковом покрове — лишайники, то эта ассоциация будет называться сосняк вересково-лишайниковый. Если в лесном фитоценозе достаточно хорошо выражен подлесок, то доминатные виды подлеска также включают в название ассоциации — ельник лещиново-кисличный. Русские названия луговых ассоциаций можно давать, поэтому же принципу. При этом по постановлению ботанического конгресса преобладающее в ассоциации растение ставят в названии на последнее место – душистоколосково-луговоовсянициевая ассоциация. Применяют также следующий способ названия ассоциаций: доминанты – относящиеся к одному ярусу, соединяют знаком « + », а к разным – знаком «—». Например, лисохвост луговой + ежа сборная — душистый колосок. Используют народные названия: бор, суборь и др. Наименование ассоциаций на латинском языке может состоять из двух слов. Например, ассоциация ельник-кисличник:Piceetumoxalidosum. Название образовано из корня латинского названия эдификатора ели —Picea (к которому прибавляют окончание «etum») и корня латинского названия субэдификатора кислицы — Oxalis (к которому прибавляют окончание «osum») (Быков, 1983). Имеется много классификаций по почвенному покрову, при описании фитоценозов особенно сложных, соблюдается строгое соподчинение единиц различного ранга. Классификация фитоценозов нужна для изучения, длительного мониторинга, проследить наличие сукцессий, для практических целей – для создания карт различных растительных территорий. Перед составлением карт проводят классификацию растительного покрова, обследование растительности, которая встречается ГЛАВА 2. ДИНАМИКА РАСТИТЕЛЬНЫХ СООБЩЕСТВ 2.1 Сезонная изменчивость фитоценозов Фитоценозы непрерывно изменяются во времени. Изменения в растительных сообществах происходят постоянно: одни параметры могут меняться в течение суток (например, интенсивность транспирации или фотосинтеза), другие меняются в течение вегетационного периода в связи с сезонным развитием растений (например, начало роста, цветения или плодоношения отдельных видов, листопад и др.). Сезонная изменчивость – это изменения фитоценоза, происходящие на протяжении года. Они обусловлены переменами условий существования растений от сезона к сезону (температура, влажность и др.). Сезонная изменчивость проявляется в изменении внешнего облика растительности и многих других особенностей фитоценоза (количественные соотношения между видами растений, структура, продуктивность и др.). Однако состав фитоценоза остается неизменным. Примером сезонной изменчивости может быть отмирание надземных органов некоторых травянистых растений осенью и отрастание их следующей весной. Сезонная изменчивость фитоценозов наблюдается в тех районах Земного шара, где условия жизни растений на протяжении года резко меняются. Там, где этого нет, сезонная изменчивость фитоценозов отсутствует (в дождевых тропических лесах). Общей закономерностью сезонной изменчивости является зависимость параметров растительного сообщества от изменения экологических факторов в течение вегетационного периода. Ритм развития растений и сообщества в целом определяется в основном температурным режимом и снабжением растений влагой. Образное описание сезонной изменчивости разнотравного пойменного ненадолго заливаемого луга около г.Вологды приводит в своем учебнике «Луговедение» А. П.Шенников (1941). В ходе его ежегодного развития в течение периода вегетации выделяются 7 стадий. Первая стадия – предвесенняя – продолжается от времени оттаивания почвы примерно до половины мая. В это время происходит позеленение луга от появляющихся новых побегов и развертывание свежих прикорневых листьев, появляются колоски лугового хвоща и бутоны обильной здесь фиалки (Viola arenaria). У многих растений, зимовавших без зеленых листьев, появляются надземные побеги с розетками листьев. В зависимости от температуры эта стадия наступает несколько раньше или позже, в связи с временем первой весенней волны тепла, и проходит быстрее или медленнее, в связи с обычной на севере волной холода (Дохман, 1973). С новым повышением температуры луг переходит во вторую стадию – ранневесеннюю. В ранневесеннюю стадию луг имеет фиалковый аспект от массы цветущей Viola arenaria. Одновременно цветет Glechorna hederacea и пылит хвощ. Формируется подсед – кустятся злаки, начинается стебление разнотравья. Замечается прирост мха (Thuidium abietinum). Эта стадия продолжается до конца мая, затягиваемая очередной волной холода. Третья стадия – разгар (середина) фенологической весны – начинается с новым, более устойчивым повышением температуры. Аспект характеризуется обилием цветущего Trollius curopaeus; луг от него желтеет. Тогда же цветут, зацветая одновременно или почти одновременно, Geum nvale. Taraxacum vulgare, Polygala amara, Alchimilla pubescens, Carum cam. Ranunculus acer и др.; луг делается очень цветистым. Это время усиленного роста травостоя, стебления низовых злаков, формирования 2–го яруса, постепенного затенения мха и приземистых растений. Продолжается до 10–15 июня. Четвертая стадия – конец весны – начало лета, т.е. переход от весны к лету. Массовое цветение Polygonum bistorta, луг от него розовый. Зацветают Trifolium pratense, Rumex haplorhizus и другие летние растения, и цветущих растений становится больше, чем в предыдущую стадию. Ранние злаки в это время заканчивают колошение (красная овсяница, луговой мятлик). Формируется 1–й (верхний) полог верховых злаков и высокого разнотравья. Эта стадия обычно коротка. Пятая стадия – разгар лета (или середина фенологического лета); начинается около 20 июня и продолжается до 15–20 июля. Начало ее зацветание низовых злаков (красной овсяницы и лугового мятлика) и вместе с ними многих других растений, в том числе массы Leucanthemum vulgare и Campanula glomerata, характеризующих пестрый аспект луга. Это время – апогей фенологического развития нашего луга. Травостой сформирован полностью. Число цветущих видов наибольшее. Зацветают и поздние злаки. В шестой стадии – конец лета – цветистость луга уменьшается, и в аспекте его более заметны окраски обильных плодоношений. Заметны также увядание и отмирание нижних листьев и побегов. Число цветущих видов уменьшается, хотя некоторые (иозднелетние) зацветают только теперь (Hieracium umbellatum, Agropyrum re–pens, Delphinium elatum). Формируются почки перезимовывания. Взамен отмирающих генеративных побегов появляются новые вегетативные. Характерно, что увядание прогрессирует несмотря на то, что погода еще теплая; лишь в конце этой стадии – около половины августа – начинаются заметные похолодания. И хотя после них снова бывает еще довольно долгое время тепло, луг переходит в осеннее состояние. Седьмая стадия – осеннее состояние луга – время явного преобладания во внешности луга признаков увядания и отмирания травостоя. Цветущие растения единичны или отсутствуют. Продолжается новое побегообразование, особенно кущение злаков, дающих новое поколение вегетативных побегов под отмирающим весенне–летним поколением плодущих побегов. Эта стадия продолжается до морозов и снега. Графическое изображение прохождения фенологических фаз видами, слагающими сообщество, называется фенологическим спектром. Обобщенной характеристикой фенологического состояния сообщества является число видов, находящихся в одной фазе развития, например цветения. Число цветущих видов постепенно увеличивается, достигает максимума в середине лета (с середины июня до середины июля) и постепенно уменьшается. Ритм развития видов индивидуален. Он отражает неодинаковое отношение к экологическим факторам. Существенную роль в сезонной динамике играет изменение световой обстановки в толще растительного покрова. В лесостепной дубраве подпологом древостоя большинство видов ускоренно накапливают фитомассу до момента полного развития листьев деревьев (рис.1), т.е. в условиях достаточной освещенности. Пик цветения наблюдается также до смыкания полога (рис.2). Значительную долю в образовании весенне–раннелетнего пика составляют эфемероиды (Scilla sibinca. Corydalis halleri, Anemone ranunculoides, Gagea lutea. G.minima. Ficana verna), требовательные к свету и способные быстро развиваться при сравнительно невысоких температурах благодаря запасенным в подземных органах питательным веществам. Пик весеннего цветения формируется и видами, цветущими несколько позднее эфемероидов, – p. Viola, Latkyrus pern us. Carex pilosa, Asarum europaeum, Glechoma hederacea и др. (Ипатов, 1997). 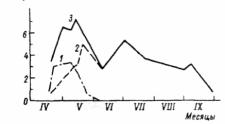 Рис.1. Динамика надземной массы в травяном покрове дубравы (липо–дубняк снытевый, лесостепь) (по: Горышина, 1969): 1 – эфемероиды, 2–виды летней синузии, 3 – суммарная фитомасса. В течение вегетационного периода часто изменяется вертикальное раcпределение листовой поверхности и фитомассы. Особенно ярко это выражено в сомкнутых крупнотравных сообществах, как, например, с господством лабазника – Ftlipendula camtschatica (рис.3). Весной листовая поверхность растений распределена по высоте сообщества более или менее р 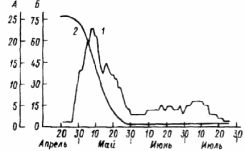 авномерно. Затем ее основная часть перемещается вверх, происходит это из–за отмирания ранневегетирующих растений и интенсивного роста крупных трав. Рис. 2. Кривая цветения травянистых видов под пологом дубравы (липо–дубняк. лесостепь) (по: Горышина, 1969): А, 1 – число цветущих видов; Б, 2–относительная освещенность (%) на уровне травяного покрова. 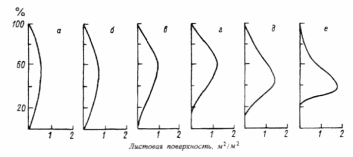 Рис. 3. Распределение листовой поверхности по высоте у лабазниковых (Filipendula camtschatica) травостоев в расчете на участок площадью 1 м2, Камчатка (по: Степанова и др., 1981): а–15 июня, 6–23 июля, в –6 июля, г – 15 июля, д–28 июля, е –9 августа. Сезонная изменчивость сообществ неодинакова. Она определяется со– ставом сообществ и особенностями изменения экологических факторов в разных климатических зонах и экотопах. В упомянутом лабазниковом со– обществе крупные травы образуют при полном своем развитии сплошной листовой полог. Под ним возникает крайний дефицит света, препятствующий произрастанию низкорослых растений. Листовая поверхность в течение сезона «поднимается» вверх. В высокотравном луговом сообществе в степной зоне с увеличением размеров растений также наблюдается такой подъем. Однако в целом листовая поверхность оказывается более или менее равномерно распределенной по высоте, поскольку здесь не образуется плотного полога, перехватывающего свет (рис. 4).  Рис. 4. Изменение вертикального распределения относительной листовой поверхности (ОЛП) в высокотравном луговом сообществе, степная зона, дно балки (по: Утехин, 1977). 1 – начало июня, 2– начало августа. В широколиственных листопадных лесах весной при обилии света под необлиственным еще древостоем развиваются синузии эфемероидов, часто образующие сплошной ковер, но к середине лета в надземной своей части полностью исчезающие. В сомкнутых вечнозеленых еловых лесах таежной зоны из–за ограничения светопого довольствия такие возможности для эфемероидов отсутствуют. Сезонная смена синузий здесь не выражена. Под пологом березовых же лесов в тайге синузия эфемероидов успевает сформироваться до полного развития листвы березы, правда видовой состав синузии беден и представлен в основном ветреницей (Апетопа nemorosa). Впрочем, в вегетативном состоянии многие особи ветреницы сохраняются в течение всего лета (Ипатов, 1997). В степи после образования основной массы в весенний и раннелетний период ее уровень мало меняется; Ограничивают образование фитомассы высокие температуры, а главное, недостаточное снабжение растений водой. На субальпийских лугах, в условиях обеспеченности влагой динамика фитомассы следует за изменением температурного режима (рис.5). 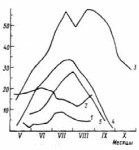 Рис. 5. Сезонное изменение зеленой массы в разных сообществах (Гордеева, 1965; Маилов, 1971; Коваленко и др., 1973): 1 – тонковатополынно–типчаково–ковылковое, 2–тонковатополыннотырсиковое (степные сообщества, Центр. Казахстан); 3–злаково–бобово–разнотравный послелесной субальпийский луг, заповедный; 4 – красно–овсяницевый луг, 5–белоусовый луг – субальпийские луга в Карпатах. Своеобразный ритм развития имеют пустынные сообщества. Во многих пустынных сообществах однолетние растения создают основные аспекты (внешний вид) и производят большую часть фитомассы. В сезонном ритме сообществ выделяются три группы видов: ранневесенние эфемеры, поздневесенние эфемеры и летние однолетники (Нечаева. Василевская, 1945). Они различаются строением, позволяющим растениям адаптироваться к режиму влажности, тепловому режиму в разное время вегетационного периода. Ранневесенние эфемеры (Ceratocephalus falcatns. Meniocus linifolius, Streptoloma desciforum) прорастают в ноябре, цветут в начале марта и засыхают в середине апреля. Они развиваются в период с достаточным количеством влаги и сравнительно невысокими для пустыни температурами и имеют мезоморфное строение. Поздневесенние эфемеры (Lappula seniglabra, Нурссоит pendulum, Malcolmia grandiflora. Isatis emarginaia, Senecio subdeniatus, Centaurea gyzensis, Delphinium camptocarpum) прорастают в декабре, цветут в апреле, засыхают к середине мая. Ксероморфные черты (утолщенная пластинка листа, столбчатая ткань с верхней и с нижней стороны) позволяют им развиваться в условиях большего дефицита влаги и более высоких температур. Продолжительность вегетации летних однолетников больше, чем у эфемеров, и проходит в другие сроки: прорастают они в марте–апреле и завершают развитие осенью (в конце сентября – начале октября). Эти виды – представители семейств Chenopodiaceae и Euphorbiaceae (Chrozophora sabulosa, Euphorbia cheirokpis, Salsola pellucida, Horamnovia uhcina. Agnophyllum latifohum, A.minus, Londesia enantha, Kochia schrenkiana. Airiplex dimorphostegia), развиваются в жестких условиях ограниченного водного довольствия и высоких температур. Устойчивость этих растений к засухе обеспечивается тем, что в их стеблях и листьях развита крупноклеточная водозапасающая ткань, в тканях накапливаются хлориды и сульфаты, благодаря этому клеточный сок имеет повышенное осмотическое давление. У растений семейства Euphorbtaceae имеются млечники, заполненные слизистыми веществами, смолами, маслами. Разумеется, в сообществах участвуют и другие виды, многолетники, как монокарпики, так и поликарпики. В пустыне сезонная динамика отличается глубокими и резкими изменениями. В описанных пустынных сообществах наблюдаются два пика вегетации: летний и осенне–весенний. В некоторых экотопах сезонный ритм может в основном зависеть от биологических особенностей самих растений. В тугайных травяных сообществах, монодоминантных, прирост фитомассы определяется ритмом развития доминирующих видов. Прирост фитомассы прекращается по достижении доминантами предельных размеров. Период же роста у разных видов неодинаков. Например, у Typha minima он заканчивается в конце июня, а у Phragmites australis продолжается непрерывно в течение вегетационного периода (Ипатов, 1997). Последовательность сезонных изменений у одного и того же сообще– ства из года в год повторяется, но сроки прохождения фаз и отдельные частности различаются. 2.2 Флуктуация Если условия произрастания видов в сообществе заметно меняются от года к году, может наблюдаться разногодичная изменчивость – флуктуации. Продолжительность цикла флуктуаций не превышает 10 лет. Например, урожайность пойменного луга в долине реки Белой в разные года изменяется в 10 раз. Флуктуации фитоценозов у северной границы степей приводят к тому, что границы степи во влажный год может сместиться на юг, а в сухой – на север. В экстремальные годы граница может сместиться на 100 км. Разногодичная изменчивость, или флуктуации, – это изменения фитоценоза от года к году. Они чаще всего связаны с тем, что условия существования растений в разные годы не одинаковы, причем иногда различаются в сильной степени (например, в некоторые годы выпадает много осадков, в другие – мало). Разногодичная изменчивость фитоценозов хорошо выражена на пойменных лугах, заливаемых весной при разливе рек. На этих лугах в разные годы сильно разрастаются и становятся доминирующими то одни растения, то другие. Количественное соотношение разных видов растений год от года сильно меняется, хотя общий видовой состав остается неизменным. Разногодичные изменения в данном случае обусловлены тем, что отдельные годы сильно различаются между собой по продолжительности весеннего паводка. В годы с продолжительным паводком сильно разрастаются более влаголюбивые растения (например, лисохвост луговой), в годы с кратковременным паводком – менее влаголюбивые (например, клевер луговой). Разногодичные изменения фитоценозов хорошо выражены и в степях. Здесь в некоторые года очень обильно цветут ковыли и степь летом напоминает седое море, которое колышется от ветра. В другие же годы ковыли цветут, напротив, довольно слабо и не дают сплошного фона. Отмеченное явление объясняется тем, что в разные годы выпадает то больше, то меньше осадков. В условиях степи на фоне общего дефицита влаги это очень важно. Если от года к году условия жизни растений меняются плавно, то разногодичная изменчивость фитоценозов выражена слабо. Это имеет место, например, в таежных хвойных лесах, где растения всегда обеспечены влагой. Фитоценозы тайги от одного года к другому меняются сравнительно мало. При разногодичных изменениях фитоценозов видовой состав растений остается прежним. Изменяются только господствующие растения, численность цветущих экземпляров и т.д. Происходят как бы колебания около определенного «среднего» уровня. Они не приводят к перестройке фитоценоза и смене его каким–либо другим. Возрастная изменчивость наблюдается только в лесных фитоценозах, причем таких, где деревья более или менее сходны по возрасту (древостой бывает одновозрастным обычно тогда, когда лес сформировался на сплошной вырубке, пожарище, заброшенной пашне и т.д.). На протяжении жизни одного поколения деревьев, образующих древостой, происходят существенные изменения в нижних ярусах лесного фитоценоза – подлеске, травяно–кустарничковом ярусе, мохово–лишайниковом покрове. Это хорошо заметно, например, в еловых лесах таежной полосы. В молодом, очень густом ельнике из–за сильного затенения на почве не развиваются никакие растения. В ельнике более старшего возраста, где света под пологом становится больше, появляется сначала моховой покров, а затем отдельные травы и кустарнички. В старом ельнике обычно хорошо развит как моховой, так и травяно–кустарничковый покров Возрастные изменения в нижних ярусах лесных фитоценозов обусловлены тем, что с возрастом леса увеличивается освещенность под пологом деревьев, ослабевает напряженность корневой конкуренции в почве за влагу и питательные вещества и т.д. Возрастные изменения происходят также и в древесном ярусе (Прокопьева, 2009). По мере увеличения возраста деревьев они становятся крупнее, число их на единице площади уменьшается и т.д. Возрастные изменения лесных фитоценозов обычно носят циклический характер. Достигнув старого возраста, древесный ярус распадается, отмирает. Образуется открытое пространство, на котором вновь начинает формироваться лесной фитоценоз. Он снова проходит в своем развитии различные возрастные стадии (Прокопьева, 2009). Сами флуктуации изменяются во времени в том смысле, что полного возврата к прошлому состоянию не происходит. Это объясняется тем, что движущая сила разногодичных изменений – погодные условия одно– временно колеблются в разных ритмах. На краткосрочные колебания накладываются долгосрочные ритмы, охватывающие большое число лет. Экотоп, в частности климат, имеет и длительные тенденции изменения, вызывающие соответствующие сдвиги в сообществах. Изменяющаяся растительность меняет и биотоп, что также приводит к направленным изменениям в сообществах. Если флуктуации трактовать как колебания параметров сообщества около средней периодической характеристики и принять во внимание, что средняя по указанным причинам имеет тренд, то становится очевидным, что полной цикличности во флуктуационной изменчивости и не может быть. Иллюстрирует высказанные соображения схема наложения всех изменений друг на друга (рис.6). 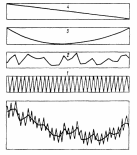 Рис.6. Схема совмещения (внизу) сезонных изменений (1) , краткосрочных флуктуации (2), длительных флуктуации (3) и направленного изменения (4) растительного сообщества. Подобная схеме картина обнаружена в исследовании В. А. Зайковой (1980). В течение 15 лет она наблюдала изменения луговой растительности в Карелии (рис.7). За этот период температура, осадки, баланс влаги, естественно, не оставались постоянными. 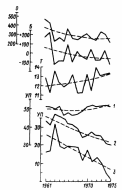 Рис. 7. Флуктуации и тенденция изменения параметров среды луговых сообществ с господством: Festuca praitnsis (1), Agrostis tennis (2), Nardus siricta (3) (Средняя тайга, Карелия) (по материалам: Зайкова, 1980): О – осадки, мм; Б – баланс влаги; X–средняя суточная температура воздуха за вегетационный период (20 мая – 30 сентября), У П – увлажнение и плодородие, оцененные по растительности, в условных единицах. Заключение Главнейшими особенностями фитоценоза являются взаимодействие между образующими его растениями, с одной стороны, и взаимодействие между растениями и окружающей средой – с другой. Влияние растений друг на друга имеет место только тогда, когда они более или менее сближены, соприкасаются своими надземными или подземными органами. Совокупность отдельно стоящих растений, не оказывающих влияния друг на друга, нельзя назвать фитоценозом (Прокопьева, 2009). Формы влияния одних растений на другие разнообразны. Они были подробно рассмотрены нами ранее. Однако не все эти формы имеют одинаковое значение в жизни растительных сообществ. Совместная жизнь растений в фитоценозе, когда они в той или иной степени влияют друг на друга, накладывает глубокий отпечаток на их внешний облик. Это особенно заметно в лесных фитоценозах. Деревья, образующие лес, сильно отличаются по внешнему виду от одиночных деревьев, которые выросли на открытом месте. В лесу деревья более или менее высокие, кроны их узкие, высоко подняты над землей. Одиночные деревья значительно ниже, кроны их широкие, низко опущенные (Василевич, 1983) Результаты влияния растений друг на друга хорошо заметны также в травяных фитоценозах, например на лугах. Здесь растения имеют меньшие размеры, чем при одиночном произрастании, менее обильно цветут и плодоносят, а некоторые и вовсе не цветут. В фитоценозах любого типа растения взаимодействуют между собой и это сказывается на их внешнем облике и жизненном состоянии (Ипатов, 1997). Взаимодействие между растениями, с одной стороны, и между ними и окружающей средой – с другой, имеет место не только в естественных растительных сообществах. Оно есть и в тех совокупностях растений, которые созданы человеком (посевы, посадки и др.). Поэтому их также относят к числу фитоценозов (Василевич, 1983). ЛИТЕРАТУРА Александрова, В. Д. Классификация растительности. Обзор принципов классификации и классификационных систем в различных геоботанических школах [Текст] / В.Д. Александрова. – Л.: Наука, 1969. – 275 с. (Александрова, 1969) Быков, Б. А. Введение в фитоценологию [Текст] / Б. А. Быков. – Алма-Ата.: Наука, 1983. – 249 с. Василевич, В. И. Очерки теоретической фитоценологии [Текст] / В.И. Василевич. – Л.: Наука, 1983. – 249 с. (Василевич, 1983) Дохман, Г. И. История геоботаники в России [Текст] / Г.И. Дохман. – М.: Наука, 1973. – 287 с. (Дохман, 1973) Ипатов, В. С. Фитоценология [Текст] / В. С. Ипатов – СПб.: Издательство С.–Петербургского университета, 1997. – 316 с. Прокопьева, Л. В. Фитоценология: учебное пособие [Текст] / Л. В. Прокопьева. – Йошкар–Ола: Map. гос. Ун–т, 2009. – 128 с. Разумовский, С.М. Закономерности динамики биоценозов [Текст] / С.М. Разумовский. – М.: Наука, 1981. – 231 с. Уиттекер, Р. Сообщества и экосистемы [Текст] / Р. Уиттекер. – М.: Прогресс, 1980. – 326 с. Шенников, А. П. Введение в геоботанику [Текст] / А. П. Шенников. – Л.: Издательство ЛУ им. А.А. Жданова, 1964. – 447 с. |