Лекция 6 Планктология. Зоопланктон Краткий очерк истории понятий
 Скачать 0.8 Mb. Скачать 0.8 Mb.
|

Вертикальные миграции зоопланктонаОрганизмы зоопланктона не могут противостоять сносу, но в толще воды, даже по ходу сноса слабым течением они постоянно перемещаются (мигрируют) по вертикали, находя свое место в температурном градиенте, уходят вглубь от яркого света, находят пищу растительную, микробную или, хищничая, одновременно спасаясь от врагов, выбирают условия для размножения, зимовки и т.д. Исследуя вертикальное распределение зоопланктона или от- дельных систематических групп в течение суток в разные сезоны года, ученые видят определенные закономерности в этом распре- делении, отражающие биологические особенности отдельных ви- дов (питание, размножение, онтогенез), популяционные отноше- ния видов, общую экологическую ситуацию в водоеме и изменение ее под влиянием хозяйственной деятельности человека: загрязнения, подогрев или изменения гидрологического режима. Мы рассмотрим ряд миграций зоопланктеров, отражающих се- зонные и суточные изменения сред обитания и биологические особенности видов. Сезонные миграцииСезонные изменения вертикального распределения планктона изучаются по биологическим сезонам: зима, весна, лето и осень. Наиболее отчетливо они выражены в высоких и умеренных широ- тах, а также в глубоких водоемах. Различают зимние, летние и круглогодичные виды зоопланк- тона. Весенний и осенний зоопланктон обычно составляют онто- генетические стадии первых, вторых и третьих. Сезонные явления в распределении зоопланктона можно про- следить на примере оз. Плещеево (Столбунова, 2006). Горизонтальное распределение зоопланктеров в озере далеко неравномерно и определяется комплексом факторов: глубиной, динамикой водных масс, температурой. Важную роль играет пи- щевой фактор и даже грунты, на поверхности которых зимуют не- которые виды циклопид, покоящиеся яйца кладоцер, коловраток. В зимний период, когда влияние гидродинамических факто- ров на водную массу ослабляется, зоопланктон распределен осо- бенно неравномерно. Его максимальное развитие наблюдается в марте. Наиболее богата центральная часть озера и участки с глу- бинами до 7 м, где летом располагается эпилимнион (до 1,05– 1,57 г/м3). Основу зоопланктона составляют холодолюбивые ко- ловратки и Eudiaptomus graciloides. После вскрытия водоема прогрев мелководий от 7,5°С до 13,4°С вызывал массовое развитие коловраток до 70 тыс. экз./м3. В глубоководных, менее прогретых участках с температурой воды у поверхности 6,8 – 8,1°С развивался в основном холодолюбивый Cyclops kolensis (до 135 тыс. экз./м3 и 2,51 г/м3). С наступлением лета, когда температура воды у поверхности достигает 21 – 24°С, возрастает роль кладоцер и летних веслоно- гих. В поверхностном слое (0 – 2 м) в центре озера Daphnia cucul-lata достигала 2,22 г/м3, Bosmina coregoni – до 1,95 г/м3, Mesocyc-lopsleuckarti – до 0,50 г/м3. В глубоководной зоне озера, а также на свале глубин образуются наиболее плотные скопления зоо- планктона (биомасса до 5 г/м3 и более). При любом из преобла- дающих направлений ветра наблюдается тенденция к увеличению биомассы зоопланктеров в двух вихревых зонах циркуляции вод в озере и разрежения плотности между ними (рис. 10). 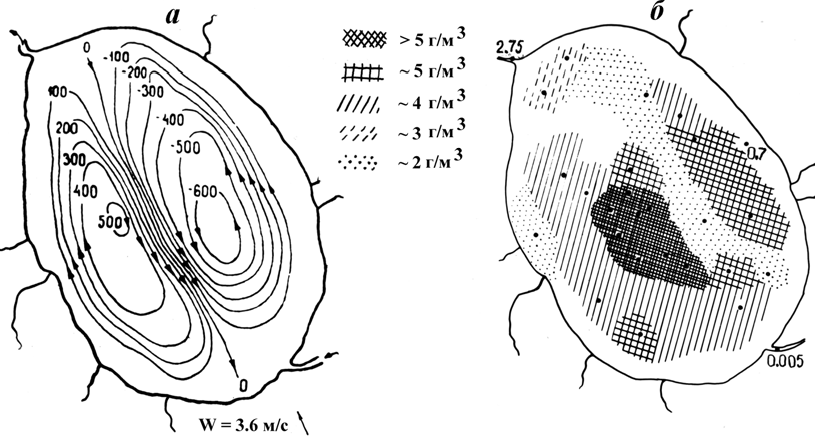 Рис. 10. Схема интегральной циркуляции вод (а, по: Поддубный,Литвинов, 1983) и горизонтальное распределение зоопланктона (б)в слое 0–2 мвлетнее время (из: В.Н.Столбунова, 2006) В осенний период с понижением температуры воды до 11 – 13°С в планктоне еще присутствуют летние Keratella quadrata, K.cochlearis, Conochilusunicornis, из ракообразных многочисленна Bosminacoregoni, встречаются Daphniacucullataи Eudiaptomusgraciloides. Распределение зоопланктона в поверхностном слое определяется погодными условиями, и при ветровом перемеши- вании воды оно более равномерно. Межгодовые колебания общей численности зоопланктона в поверхностном слое пелагиали озера за вегетационный период мало отражались на величине биомассы, так как основу последней составляли ракообразные, за исключением 1989 г., когда преобла- дала крупная Asplanchna priodonta(табл. 1). Таблица 1 Средние количественные показатели зоопланктона в поверхностном слое 0 – 2 м профундали оз. Плещеево за вегетационный период в разные годы
Вертикальное распределение зоопланктона в озере изменя- ется в зависимости от условий сезона года, прежде всего от тер- мического режима и связанной с ним стратификации водной мас- сы, а также от немаловажного пищевого фактора. В зимний период при устойчивой стратификации водной тол- щи и температуре воды 0 – 2,9°С, в поверхностном слое наиболее многочисленны коловратки родов Keratella, Polyarthra, Synchaeta; здесь максимальное содержание кислорода, минимальная тем- пература, присутствуют бактерии и водоросли.Придонные ско- пления образуют Filinia maior и Conochiloides natans, хорошо пе- реносящие дефицит кислорода, а также Asplanchna priodonta. В некоторые годы придонные скопления образовывал эвритермный Conochilus unicornis. Eudiaptomus graciloides заселяет всю толщу пелагиали озера с тенденцией возрастания численности к поверх- ности. Его количество значительно больше, чем Cyclops kolensis, который образует небольшие скопления в придонных слоях. Вет- вистоусые Bosmina coregoni, Daphnia longispina и D. cristata сосре- доточены в металимнионе и придонных слоях. Весной, в период гомотермии, при температуре 5–6°С коло- вратки и ракообразные распределяются по вертикали относитель- но равномерно. В конце мая, когда температура воды повышается до 17,0 – 17,5°С, начинает формироваться слой температурного скачка. В период летней стратификации водной толщи вертикаль- ное распределение популяций зоопланктона позволяет выделить два основных комплекса: тепловодный многокомпонентный (Ke-ratella quadrata, K. cochlearis, Pompholyx sulcata, Conochilus un-icornis, Daphniacucullata, Bosminacoregoni, Eudiaptomusgraci-loidesи др.) и холодноводный малокомпонентный (Keratellahiemalis, Filinia maior, Daphnia cristata). Наибольшей плотности зоопланктон достигает в эпилимнио- не, где располагается оксиклин и интенсивно идут первичные продукционные процессы. По мере охлаждения озера, ускорения циркуляции вод и значительном ветровом перемешивании водной толщи наступает осенняя гомотермия. Распределение коловраток и ракообразных по вертикали становится более равномерным. Суточные миграцииСуточные миграции зоопланктеров – интереснейшая научная и практическая тема исследований. Ни одна серьёзная работа по зоопланктону, например диссертация, не обходит эту тему. Здесь изучаются разные стороны биологии и экологии видов, трофиче- ские связи и отношения популяций. Суточные миграции происхо- дят во всем Мировом океане и практически во всех озерах и водо- хранилищах. Трудно себе представить, но это происходит, что триллионы тонн планктонных организмов каждые сутки подни- маются из глубин к поверхности, питаются там, размножаются и опускаются обратно. Суточные миграции могут быть горизон- тальными и вертикальными. Примерами горизонтальных миграций могут быть суточные перемещения Polyphemus pediculus из разрозненных групп в ноч- ное время в локальные плотные скопления по мере увеличения освещенности с восходом солнца. К вечеру скопления рассредо- тачиваются. Подобные скопления можно наблюдать у Daphniamagna, но в затененных водоемах в дневное время. При ярком дневном освещении дафнии уходят на глубину. Изучение вертикальных миграций зоопланктона предполагает экологическую оценку этого грандиозного явления, вероятно дей- ствующего со времени зарождения жизни в толще вод океана. Зада- чи изучения в основном сводятся к установлению причин, времени и глубины нахождения видов, групп (Cladocera, Copepoda, Rotatoria) и их представителей на определенных стадиях развития и размно- жения. Наблюдения (отбор проб) проводятся круглосуточно, через 2 – 3 часа, на глубинах от поверхности до дна, обычно через 2 м. Наиболее полные исследования в этом плане проводятся во все 4 биологических сезона и обычно в открытой, глубоководной части озера или водохранилища. В морях это обычно специальные эпизо- дические исследования из-за их трудоемкости и дороговизны. Существует целая история изучения вертикальных миграций. Первые исследования суточных вертикальных миграций ра- кообразных были проведены на оз. Леман в 1874 г. (Forel, 1874, 1879) и на оз. Боденском (Weismann, 1877), где наблюдались су- точные миграции Leptodora hyalina(L. kindtii). Уже к началу ХХ в. были установлены некоторые закономер- ности суточных вертикальных миграций зоопланктона. Эти зако- номерности касались интенсивности и размаха (амплитуды) ми- граций у разных видов, неодинакового поведения одних и тех же видов в разных водоемах, изменения характера миграций в связи с возрастом (стадией), с полом, в зависимости от сезона года, изме- нения времени подъема к поверхности и ухода вглубь у разных видов. Делались тогда же и первые попытки истолковать это яв- ление, установить факторы, управляющие им. Кушинг (Gushing, 1951) изучал миграции ракообразных в све- товом градиенте, когда рачки находили свой световой оптимум, где они могут успешно питаться. Величина светового оптимума неодинакова у разных видов. В связи с этим время появления зоопланктеров у поверхности и опускания их на глубину у разных видов может быть весьма не- одинаковым. Различают мигрантов сумеречных и ночных. Первые концентрируются у поверхности в большом количестве вечером и на рассвете, вторые – ночью, количество первых у поверхности падает днем и ночью, а у вторых – днем. Оказалось, мелководные виды ведут себя как послеполуден- ные мигранты, более глубинные – как вечерние, а еще более глу- боководные – как ночные мигранты. При этом чем глубже вид живет в озере, тем сильнее варьирует его средняя дневная глубина от случая к случаю и тем больше вертикальная протяженность его популяции. Световой оптимум различен у разных стадий, полов и даже генераций одного вида, может зависеть от физиологического состояния организма. Отсутствие вертикальных суточных миграций отмечено в олиготрофных гумифицированных озерах с коричневой водой и незначительной прозрачностью из-за довольно однородных усло- вий во всей толще воды в течение суток. Хатчинсон (Hutchinson, 1967) различает три типа ночных ми- граций: Подъем начинается перед или вскоре после захода солнца, и животные достигают поверхности незадолго до полуночи, спуск начинается рано утром, как только небо начинает светлеть (юве- нильные Daphnia longispina в оз. Лох-Дерг, копеподы в оз. Лю- церн, Diaphanosomaв оз. Титизее). Подъем продолжается всю ночь, и максимум у поверхно- сти отмечается перед рассветом или в умеренных широтах ле- том – около 4 ч утра Eudiaptomus gracilis и Eurytemora velox в оз. Люцерн). Нет хорошо выраженного ночного подъема, скорее, имеет место ночное опускание, длящееся всю ночь (Cyclopsstrenuusв оз. Нозири и Macrohectopus branickii в оз. Байкал). При обзоре поведения родов, видов или одного и того же вида в различные периоды его жизни, в разные сезоны у разных стадий и даже генераций или в разных водоемах обнаруживается удиви- тельное разнообразие суточного цикла с уклонением от общей схемы и даже аномалии в поведении. Наблюдения показали, что наиболее активно мигрируют взрослые рачки. Молодь (науплии) мигрируют мало и в основном находятся в слое воды, содержащем больше растительного детри- та и разлагающих его микроорганизмов, которые составляют ос- новную пищу науплий и копеподитов. Этим слоем часто оказыва- ется слой температурного скачка. Онтогенетические миграцииБольшая часть пелагических животных, особенно населяю- щих промежуточные и глубинные воды океана, меняют глубину своего обитания на разных стадиях развития. Принципиальная схема возрастных миграций у всех планктонных животных, за не- многими исключениями, однотипна. Половозрелые формы, или во всяком случае половозрелые самки, преобладают обычно на наи- больших глубинах обитания вида и там размножаются. Личинки (или яйца) поднимаются на наименьшие доступные виду глубины. Реже для икрометания поднимаются самки. В обоих случаях ли- чинки откармливаются в богатых пищей верхних слоях и лишь по мере роста опускаются на большие, бедные пищей, но зато менее населенные глубины. Амплитуда возрастных миграций определя- ет степень эврибатности вида и может колебаться от нескольких десятков и сотен до нескольких тысяч метров. Естественно, указанным образом распределяется не вся попу- ляция, так как у многих глубоководных животных эврибатная мо- лодь встречается по всей глубине обитания вида и лишь преобла- дает в верхних горизонтах; также и половозрелые особи многих видов поднимаются значительно выше глубины своего основного обитания (Бирштейн, 1951; Виноградов, 1955, 1959). У видов, поднимающихся в поверхностную зону, каждая фаза онтогенетических миграций (во всяком случае в средних и высо- ких широтах) приурочена к определенному времени года, и они в равной степени могут быть названы сезонными миграциями. Различные генерации одного и того же вида могут мигриро- вать по-разному и в разное время. Среди интерзональных видов, диапазон обитания которых дос- таточно широк, в зависимости от того, где происходит икрометание (в поверхностной зоне или в глубже лежащих водах), можно выде- лить два типа онтогенетических миграций. В одном случае в по- верхностные слои для икрометания поднимаются взрослые формы, в другом – молодь, вылупившаяся в глубинных слоях. В морях Северной Атлантики первым типом миграции обла- дают наиболее многочисленные верхне-, интерзональные виды копепод Calanus finmarchicus(Мантейфель, 1941). Причины миграцийБолее чем столетние исследования миграций зоопланктеров показали несколько разных причин, неодинаковых для водоемов разного типа, широтного расположения, разного светового режи- ма и особенно температурного. Объединяющей причиной может быть нахождение организмами лучших условий для парения и пи- тания. Организмы с менее активным движением, могут переме- щаться с вертикальными токами воды в течение суток, при смене дневных и ночных температур. Ракообразные с более активным движением могут самостоятельно стремиться в слои воды, верх- ние, где они находят себе пищу и одновременно становятся неви- димыми для врагов, например, миграции морской каляниды Kala-nusfinmarchicusв Баренцевом море на сотни метров ночью к поверхности моря и уход вглубь утром. То же известно для рачка- эпишуры в оз. Байкал в летний период. Известны барические миграции планктона в морях: при по- нижении атмосферного давления перед штормами, организмы – медузы, гребневики, крылоногие и голожаберные моллюски и да- же нектонные головоногие моллюски – уходят на глубину. В озе- рах наблюдается то же. При волнении рачки и коловратки уходят из поверхностного, более гидродинамичного слоя. Здесь может быть и другая причина: даже при ряби на поверхности усиливает- ся освещенность верхнего слоя воды, и организмы уходят глубже в зону светового оптимума. Поскольку вертикальные миграции следует считать адаптив- ными явлениями, то они имеют определенную, эволюционно за- крепленную пользу для организмов. Биологические выгоды, которые животные получают от вер- тикальных миграций, следующие (Виноградов, 1968): Защита от «зрительных» планктофагов (Кожов, 1947; Ни- колаев, 1952; Мантейфель, 1960, 1961). Сохранение и расширение ареала благодаря попаданию в слои с течениями разной скорости и направления (Mackintosh, 1937; Hardy, 1953). Уход от коротковолновых (ультрафиолетовых) лучей. Избегание неблагоприятных условий в поверхностных слоях, особенно зимой, когда эти слои лишены пищи, и уход на глубины, где эти виды не питаются и живут за счет запасов собственного жи- ра и спасаются от выедания поверхностными планктофагами (Conover, 1962, 1964). Обмен генетическим материалом при перемешивании раз- личных частей популяций, приобретавших благодаря пространст- венной изоляции (разобщению) некоторые генетические различия (David, 1961). В нормально слоистой воде вертикальные миграции не только не приводят к дополнительной трате энергии, но и дают энергетическую выгоду мигрантам (McLaren, 1963). Именно энер- гетический выигрыш, получаемый при вертикальных миграциях, служит адаптивной основой их существования и широкого рас- пространения. Приходится признать (Грезе, 1969), что применять универ- сальный принцип объяснения всего многообразия вертикальных миграций не представляется возможным; в разных случаях раз- личные причины могут иметь решающую роль в определении биологической целесообразности миграций. Вертикальные ми- грации пелагических организмов полифункциональны по своему значению, представляют собой интегральные реакции, опреде- ляющиеся очень многими биологическими слагаемыми, и вырабо- таны исторически в качестве одной из адаптации видов к услови- ям своего существования. Изучение вертикальных миграций может быть самостоятель- ным или вторичным при изучении вертикального распределения зоопланктона в разные сезоны, сутки и в связи с рядом факторов: гидродинамики, температурной стратификации (прямой, обратной и гомотермии), послойной освещенности воды, газового режима др. Горизонтальное распределение зоопланктонаРаспределение планктона по акватории водоема зависит от многих факторов: площади водоема и удаленности центральной части (плеса) от берегов, морфологии озера (изрезанности берего- вой линии, а с ней и образование заливов, пойменных водоемов, эстуариев), глубины, зарастаемости прибрежной зоны, ветрового и глубинного переноса водных масс и др. Наиболее выражено го- ризонтальное распределение планктона по сезонам года (биологи- ческим сезонам). Наиболее разнообразен зоопланктон в заливах и в прибрежных зонах с развитой водной растительностью. Разнообразие убывает от периферии к центру. Размерный состав зоопланктона увеличивается от центра к периферии. Одни и те же виды крупнее в прибрежной зоне, особенно в зарослях. Это связано скорее всего с гидродинами- кой и лучшими условиями питания, например, кладоцер. Горизонтальные перемещения планктона в массовом масшта- бе обычно связаны с ветровым волнением, создающим сгонно- нагонные явления. От весны к лету планктон распространяется к центру озера или водохранилища, к осени наблюдается обратная картина. Откладка зимующих яиц у Cladocera происходит на ма- лых глубинах. Из горизонтальных миграций хорошо известны и наблюдаются у Polyphemus pediculus. С увеличением освещенности воды рачки начинают смещаться к берегу, образуя на малых глубинах большие скопления. Чем ярче солнце, тем крупнее и плотнее стая. Образова- ние стаи способствует защите от хищников и создает лучшие усло- вия питания – каннибализма взрослых рачков. Такие явления скоп- ления рачков известны и для дафний. Daphniapulexможет образовывать крупные скопления у поверхности воды, но в более пасмурные дни и в затененных водоемах (лесных). В реках гори- зонтальные перемещения планктонных организмов, голопланктона и меропланктона вместе с донными мелкими организмами, чаще личинками насекомых, происходит в дрифте по течению. |