липиды. Лекция Липиды
 Скачать 1.39 Mb. Скачать 1.39 Mb.
|

Лекция 4. ЛипидыБарьерные функции липидов И в клетке, и снаружи клетки чаще всего водная среда. Возникает проблема разделения структур и веществ, которые должны или не должны находиться в клетке. Поэтому необходим барьер. В теории таким барьером может быть любое вещество, образующее плотный слой. Но на практике это не так, поскольку необходимо, чтобы прохождение веществ через барьер было контролируемым. Этого не добиться, если использовать, например, структуры из глобулярных белков или длинных тяжей полисахаридов. Для создания барьера необходимы гидрофобные соединения, такие как липиды. Барьер также должен быть непрерывным. Есть организмы, у которых определённые покровы образованы белками. Например, у трипаносом есть ундулирующая мембрана, которая представляет из себя белковый чехол, покрывающий клетку. Под этим чехлом находится жгутик, и когда он вращается, он оттягивает и деформирует мембрану, за счёт чего он передвигается. Белковая оболочка здесь служит для защиты. Также интересно, что это чехол меняется со временем. Таким образом, хозяин не успевает выработать иммунитет. В некоторых случаях белковая оболочка находится под мембраной, помогая клетке отделиться от среды. Так работает пелликула у эвгленовых. У них пелликула представляет из себя белковую ленту, закрученную в спираль по длине всей клетки. Но при этом мембрана всё равно работает, именно как химический барьер. Это – основная функция липидов. Есть организмы, у которых присутствуют только мембранообразующие липиды. Липиды – очень разнородная группа. К липидам обычно относятся вещества биологического происхождения, нерастворимые в воде и растворимые в полярных растворителях (хлороформ, бензол). При этом важно соблюдение всех трёх обозначенных условий Для того, чтобы выделить липидную фракцию из живого организма, необходимо разрушить этот организм, затем использовать воду и определённый неполярный растворитель (чаще всего используется реагент Фолча) в определённых соотношениях. После встряхивания и перемешивания этой смеси образуются четыре различных фазы: нерастворимая, в которой находятся клеточные стенки, тканевые волокна, водная фаза с водорастворимыми компонентами, фаза полярного растворителя, в котором содержатся липиды, и интерфаза между водой и растворителем, в которой содержатся промежуточные соединения. Но в липидная фракция в данном случае оказывается большое количество различных по своей химической природе соединений. Выделяют до 40 различных классов липидов. В программе курса будут рассмотрены наиболее важные из них. Другие функции липидов Липиды служат не только как образователи мембран. Прежде всего. они служат хорошим веществом для запасания: они нерастворимы, не повышают осмотическое давление, но при этом требуют кислорода для окисления и использования в качестве энергии. Липиды запасаются большинством животных и многими растениями. Но организмы, живущие в анаэробных условиях, не запасают липиды, поскольку в бескислородной среде окисление липидов невозможно. А значит, запасать их бессмысленно. Такое наблюдается у анаэробных микроорганизмов, а также некоторых эндопаразитов, живущих в бескислородной среде. В сложных многоклеточных организмах животных липиды также участвуют в транспорте веществ. У человека в крови циркулируют особые белково-липидные комплексы, переносящие липиды между различными частями организма. Прежде всего, это липиды из переваренной пищи или компоненты, требующие перераспределения или утилизации. Также необходимо отметить, что ряд липидов используются как сигнальные молекулы. Иными словами, такие липиды являются гормонами. Также некоторые липиды (в частности, воска) выполняют водоудерживающую и защитную функцию. Гидролиз и жирные кислоты Липиды делятся на две группы по своему поведению в ходе гидролиза. Некоторые жирные кислоты не подвергаются гидролизу вообще. У других же некоторые связи расщепятся, в частности сложноэфирные и реже амидные. В результате такого расщепления выделяются жирные кислоты и спирты. Так как липиды лучше гидролизуются в щелочной среде, образуются соли жирных кислот, называемые мылами. В кислых растворах образуются недиссоциирующая кислотная форма, которая выпадает в осадок. Ионы жирных кислот образую мицеллы в растворе, но они обычно небольшие. Обычное бытовое мыло часто представляет из себя натриевую соль смеси пальмитиновой и стеариновой кислоты. Именно эти кислоты, потому что источником для изготовления мыла служат отходы мясного производства. Жирные кислоты могут быть разной длины, но в любом случае у них есть одна карбоксильная группа на конце и длинный углеводородный радикал. Такая структура часто всего состоит из чётного количества атомов углерода, что связано с особым синтезом жирных кислот. По сути, жирные кислоты представляют из себя восстановленные полимеры уксусной кислоты. Жирные кислоты с нечётным количеством атомов углерода могут появляться в результате дополнительных модификаций. Цепь также чаще всего линейная, хотя разветвлённые тоже иногда встречаются. По длине цепи присутствует большое разнообразие, но преобладают жирные кислоты с длиной цепи 16 (пальмитиновая) и 18 атомов (стеариновая) (табл. 4.1). Лауриновая (С12) и маристиновая (С14) встречаются в липидах крайне редко, но их и более коротких жирных кислот много в молочном жире. Жир, входящий в состав мембраны или участвующий в запасании веществ, обладает большей длиной цепи. Более длинные цепи бывают в особых случаях. В частности, в оболочках нервных клеток. При нормальных физиологических температурах большинство жирных кислот являются твёрдыми телами 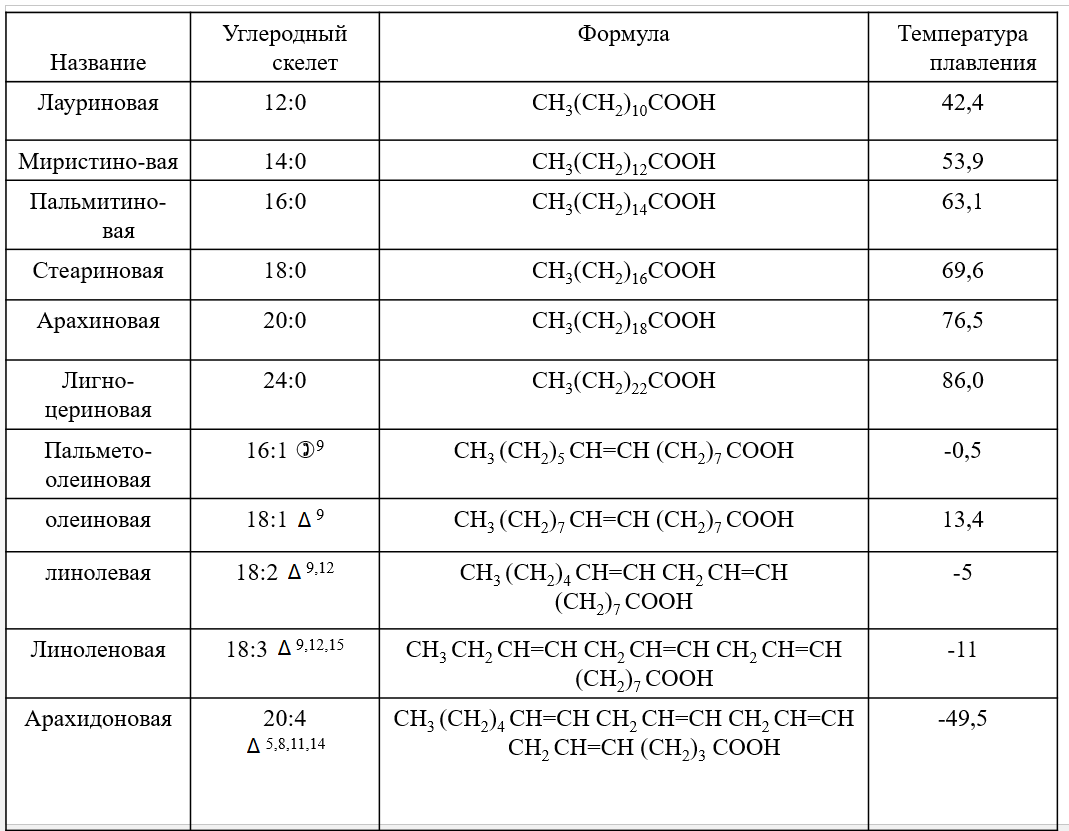 Табл.4.1.Характеристикараспространённыхжирныхкислот Упаковка жирных кислот. Насыщенные и ненасыщенные жирные кислоты Жирные кислоты по содержанию в них двойной связи, делятся на насыщенные и ненасыщенные. Обычно в ненасыщенных жирных кислотах двойные связи не сопряжены, если их несколько. Молекулы ненасыщенных жирных кислот имеют жесткий изгиб в месте, где располагается двойная связь (рис. 4.1). 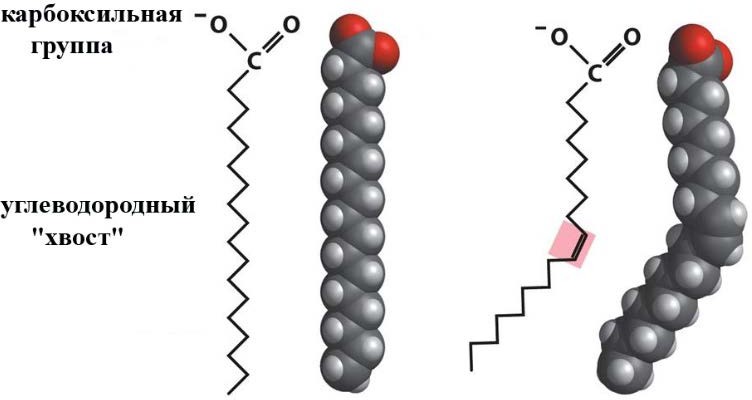 Рис. 4.1. Строение и пространственная конфигурация насыщенной стеариновой(слева)и ненасыщенной олеиновой(справа). С энергетической точки зрения, прямым и подвижным насыщенным жирным кислотам гораздо проще вытянуться и соединиться между собой за счёт Ван-дер- Ваальсовых взаимодействий по всей длине молекулы. Получается стабильная устойчивая структура (рис. 4.2). Молекулы ненасыщенных жирных кислот из-за изгиба не могут компактизироваться так же плотно, как насыщенные. Поэтому чистые ненасыщенные жирные кислоты имеют меньшую температуру плавления и в норме пребывают в жидком состоянии. 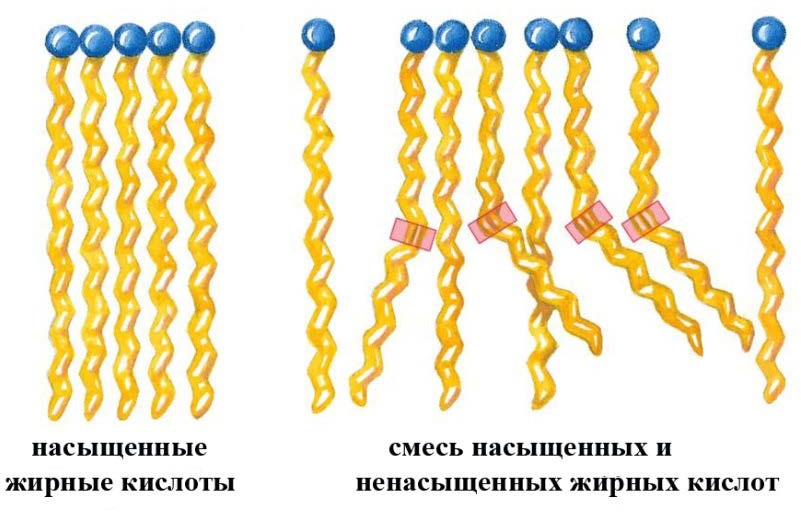 Рис. 4.2. Компактизация насыщенных и смеси насыщенных ненасыщенных жирныхкислот. На агрегатное состояние жирных кислот влияет не только количество насыщенных и ненасыщенных компонентов в составе, но ещё и природа катиона. Соли жирных кислот и калия, как правило, жидкие. Соли натрия же чаще твёрдые. Методика сокращенного обозначения названия жирных кислот также приведена в табл. 2. Первая цифра указывает на количество углеродов в цепи, вторая цифра показывает количество двойных связей. Далее цифры после знака «Δ» показывают, при каких атомах углерода есть двойная связь. Соответствующие жирные кислоты имеют также свои тривиальные названия. Ферментная система животных не может осуществить образование второй и третьей двойной связи в ненасыщенных жирных кислотах, но при этом могут образовывать все остальные. Поэтому ди- и триненасыщенные жирные кислоты для животных являются незаменимыми. Например, арахидоновая кислота у животных синтезируется из линолевой кислоты растений. Существуют и более длинные насыщенные и ненасыщенные жирные кислоты. Они встречаются реже, но они также важны для образования систем проведения нервного импульса. К чему присоединяются жирные кислоты. Триглицериды Соединения, к которым присоединяются жирные кислоты, довольно многообразны. Но в большинстве живых организмов в основном жирные кислоты соединены с глицерином. Глицериды образуются за счёт этерификации ОН-групп спирта кислотными остатками жирных кислот. 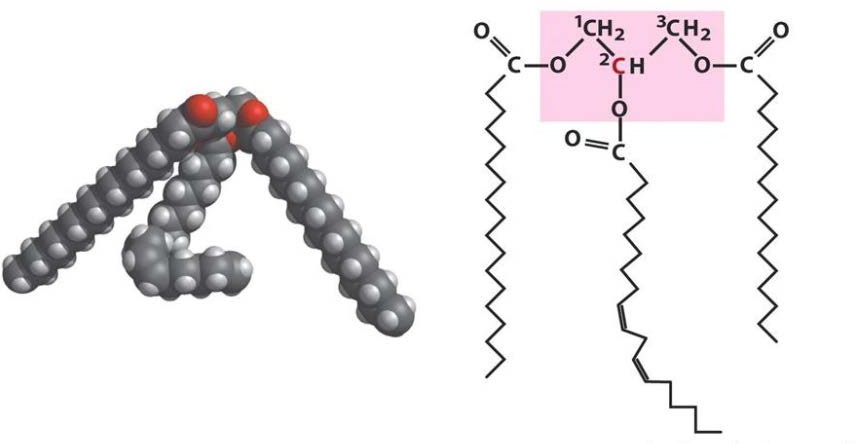 Наиболее простым является случай, когда этерификации подвергаются все три спиртовых группировки (рис. 4.3). Как правило, образуется смесь из разных жирных кислот, присоединенных к глицерину и формируется гетерогенная группа соединений. Происходит взаимодействие между гидрофобными хвостами с образованием специфической укладки. Ненасыщенные жирные кислоты создают изломы и нарушают структуру. Если много насыщенных структур, укладка более плотная. Поэтому триглицериды, в которых больше ненасыщенных жирных кислот, жидкие, а те, в которых больше насыщенных жирных кислот, твёрдые. Наиболее простым является случай, когда этерификации подвергаются все три спиртовых группировки (рис. 4.3). Как правило, образуется смесь из разных жирных кислот, присоединенных к глицерину и формируется гетерогенная группа соединений. Происходит взаимодействие между гидрофобными хвостами с образованием специфической укладки. Ненасыщенные жирные кислоты создают изломы и нарушают структуру. Если много насыщенных структур, укладка более плотная. Поэтому триглицериды, в которых больше ненасыщенных жирных кислот, жидкие, а те, в которых больше насыщенных жирных кислот, твёрдые.Рис.4.3.Смешанныйтриацилглицерид1-стеароил-2-линолеил-3-стеароилглицерин. Животные не умеют делать много двойных связей. Значит, у них преобладают твёрдые триглицериды. Их принято называть жирами. У растений же преобладают жидкие триацилглицериды, называемые маслами. Чаще всего одна единственная ненасыщенная жирная кислота находится во втором положении. Когда к глицерину начинают присоединяться различные жирны кислоты, второй атом становится асимметричным. В теории возможно образование двух изомеров. Но преобладает L-конфигурация глицерина. При запасании жидких запасных липидов возникают проблемы, связанные с их текучестью. В растения жировые капли всегда окружены снаружи мембраной. Однако, и у растений, и у животных возможно образование как твёрдых, так и жидких триацилглицеридов. Это зависит от потребностей организма. У океанических холодолюбивых животных накапливаются жидкие триглицериды, поскольку они получают много полиненасыщенных жирных кислот с фитопланктоном. Жидкие триглицериды образуют так называемый «рыбий жир». У некоторых тропических растений накапливаются твёрдые глицериды, поскольку в жарком климате они более стабильно себя ведут и меньше подвержены переходу в жидкое состояние. Классический пример – пальмовое масло. При промышленном вытапливании пальмового масла от высоких температур часть двойных связей в ненасыщенных жирных кислотах переходят в транс-состояние. В транс-форме они становятся конкурентами нормальных жирных кислот, связываясь с ферментами и транспортными системами, мешая им нормально работать. Это может привести к отравлению организма и даже раковому перерождению клеток. При этом в каждом отдельном масле или жире в состав входят совершенно разные триглицериды, как насыщенные, так и ненасыщенные (рис. 4.4) 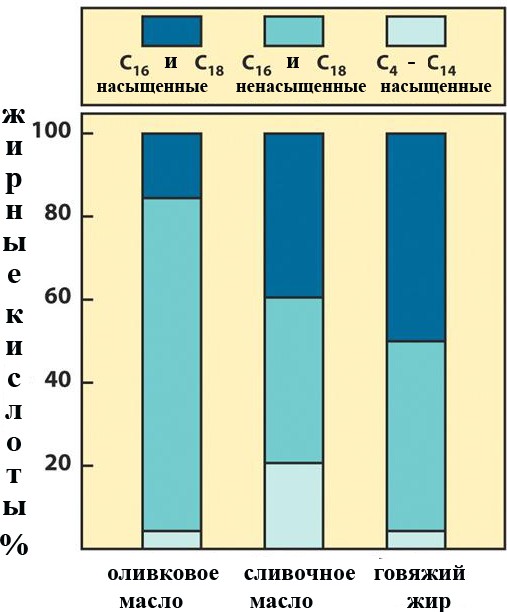 Рис.4.4.Составразличныхисточниковтриглицеридов Фосфоглицериды 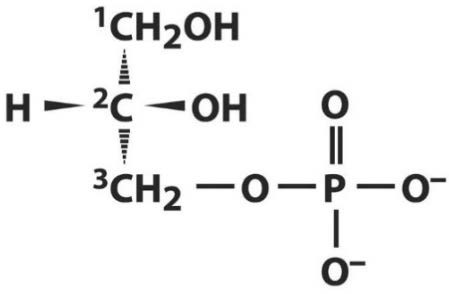 Фосфоглицериды по структуре похожи на триглицериды. Но одна из концевых гидроксильных групп этерифицируется фосфорной кислотой. При этом синтез фосфоглицеридов начинается именно с образования глицеринфосфата (рис. 4.5). В получившихся соединениях одна фосфатная и две жирнокислотных группы. К фосфату же могут присоединяться дополнительные группы. При этом в глицерине в таком случае всегда L-конфигурация. Фосфоглицериды по структуре похожи на триглицериды. Но одна из концевых гидроксильных групп этерифицируется фосфорной кислотой. При этом синтез фосфоглицеридов начинается именно с образования глицеринфосфата (рис. 4.5). В получившихся соединениях одна фосфатная и две жирнокислотных группы. К фосфату же могут присоединяться дополнительные группы. При этом в глицерине в таком случае всегда L-конфигурация.Рис.4.5.L-глицерин-3-фосфат. Как правило, ненасыщенные жирные кислоты присоединяются к С2-атому глицерина, а к С3-атому присоединяется насыщенная жирная кислота. Заместитель, присоединяющийся к фосфатной группе, зачастую имеет ярко выраженные гидрофильные свойства и иногда даже заряд (рис. 4.6). Он может иметь различную химическую природу, но в нём всегда будет спиртовая группа, за счёт которой образуется сложноэфирная связь. 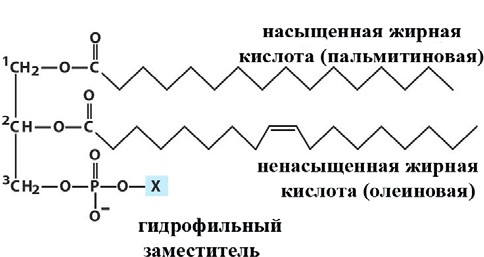 Рис.4.6.Общаяструктурафосфоглицерида. У растений в чаще используются полиненасыщенные жирные кислоты и олеиновая кислота в качестве насыщенной. Образование второй и третьей двойной связи в полиненасыщенных жирных кислотах происходит чаще всего уже после сборки фосфоглицериновой основы. Гидрофильная часть бывает достаточно большой. Возникает молекула с длинным и широким гидрофобным хвостом и гидрофильную голову. Наиболее часто встречающиеся варианты заместителей приведены в табл. 4.1. 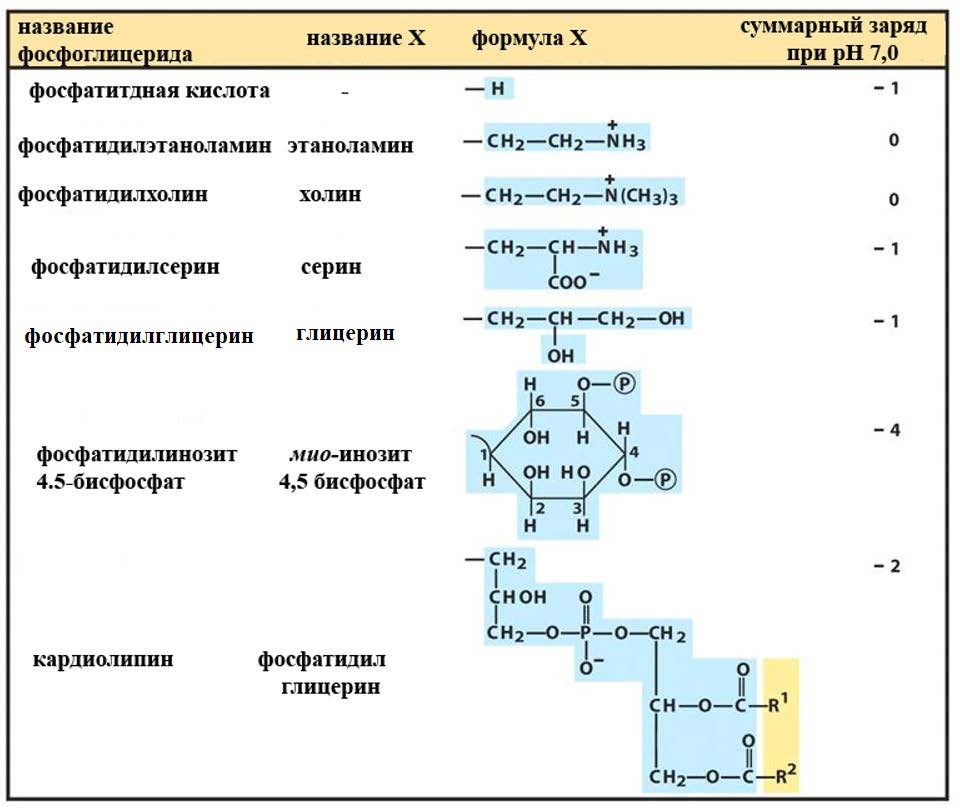 Табл.4.1.Различныевариантыгидрофильныхзаместителей. Самый простой вариант – фосфатидная кислота. В чистом виде её мало, она используется как промежуточный продукт при синтезе триглицеридов и других фосфоглицеридов. К фосфатной группе может присоединяться серин с образованием фосфатидилсерина. Возникает суммарный отрицательный заряд молекулы. Фосфатидилхолин и фосфатидилэтаноламин (кефалин) будут нейтрально заряженными молекулами, поскольку существующие заряды компенсируют друг друга. Они и фосфатидилсерин связаны друг с другом метаболически и могут служить основой для синтеза друг друга. Возможны варианты комбинации присоединения углеводного компонента или многоатомных спиртов. миоинозит может присоединяться по ОН-группе и может в дальнейшем фосфорилироваться. Фосфатидилинозитолы участвуют в регуляторных процессах. Каждый присоединённый фосфат даёт отрицательный заряд молекуле. Может также присоединяться глицерин. А к остатку глицерина в некоторых случаях может присоединяться вторая фосфатидная кислота. Образуется сложный фосфолипид, называемый кардиолипин. Кардиолипин есть в эукариотических клетках только как компонент внутренней мембраны митохондрий. Но при этом кардиолипин встречается в составе клеточной мембраны аэробных бактерий. Этот факт послужил одним из аргументов в пользу симбиотической теории происхождения эукариотической клетки. Однако, пути синтеза митохондриального и бактериального кардиолипина различаются. Кардиолипин зачастую образует комплексы с белковыми компонентами мембраны. Другие производные глицерина 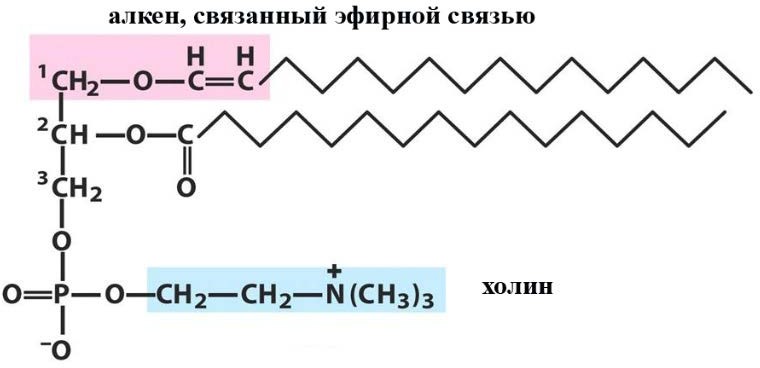 Помимо триглицеридов и фосфоглицеридов существуют другие производные глицерина. В мембранах нервных клеток часто обнаруживается плазмалоген (рис. 4.6). У него второго положения жирная кислота, а в первом положении алкеновый спирт, связанный эфирной связью с глицерином. Образуется оно из жирной кислоты, которая подвергается модификации. Это соединение при полном гидролизе образует холин, фосфорную кислоту, обычную жирную кислоту, альдегид и глицерин Помимо триглицеридов и фосфоглицеридов существуют другие производные глицерина. В мембранах нервных клеток часто обнаруживается плазмалоген (рис. 4.6). У него второго положения жирная кислота, а в первом положении алкеновый спирт, связанный эфирной связью с глицерином. Образуется оно из жирной кислоты, которая подвергается модификации. Это соединение при полном гидролизе образует холин, фосфорную кислоту, обычную жирную кислоту, альдегид и глицеринРис.4.6.Плазмалоген. По тому же принципу построен фактор активации тромбоцитов (рис. 4.7). Вместо жирной кислоты во втором положении находится уксусная кислота, а вместо жирной кислоты в крайнем положении алкан. Здесь при гидролизе вместо альдегида будет образовываться многоатомный спирт. 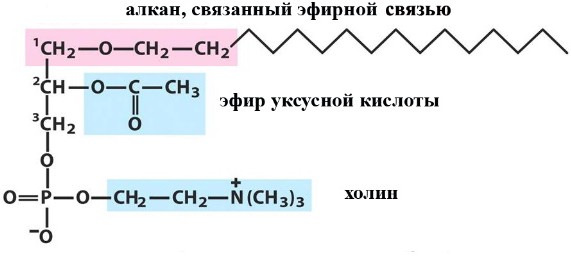 Рис.4.7.Факторактивациитромбоцитов. Существуют липиды, в которых отсутствует фосфатная группа у глицерина. Третий гидроксил глицерина образует гликозидную связь с углеводными остатками. В случае на рис. 4.8 присоединён остаток галактозы. Такого рода липиды широко распространены у растений в мембранах хлоропластов. Остаток галактозы может оказаться не один. К нему может присоединиться следующий остаток (рис. 4.9), иногда больше. Такие липиды получаются не менее гидрофильные, чем обычные фосфолипиды. Растения часто живут в условиях дефицита фосфора, а использование липидов без фосфата позволят экономить фосфор. Моносахариды, входящие в состав таких липидов, могут быть дополнительно модифицированы (рис. 4.10). 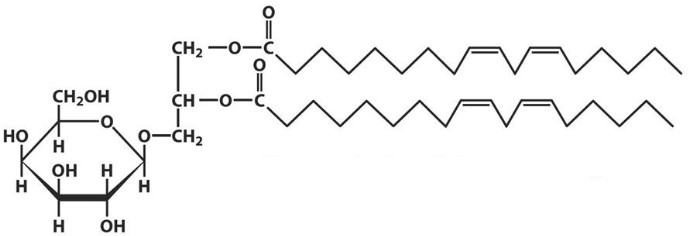 Рис.4.8.Моногалактозилдиацилглицерин. 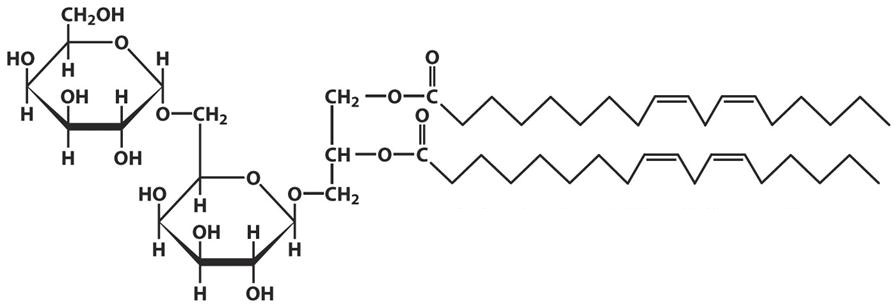 Рис.4.9.Дигалактозилдиацилглицерин. 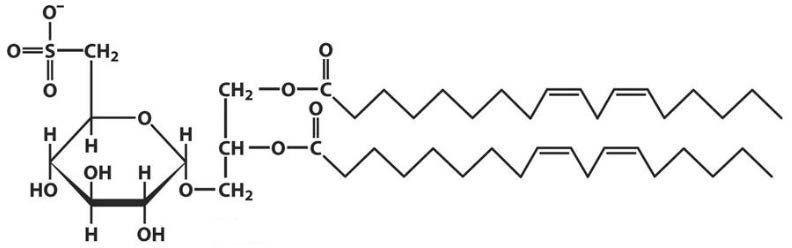 Воска Рис.4.10.6-сульфо-6-дезокси-глюкопиранозилдиацилглицерин Не всегда липиды построены на основе глицерина. Иногда используются другие спирты. К таким соединениям относятся воска. Воска широко распространены, обладают простым строением и обладают свойствами, отличными от таковых у триглицеридов. Воск состоит из остатка жирной кислоты и остатка первичного длинного спирта (рис. 4.11). У пчёл такой спирт образуется специально в соответствующих железах. Получается длинная гидрофобная насыщенная цепь, образующая прочные структуры. 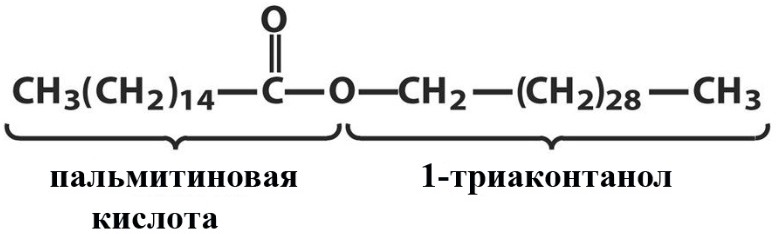 Рис.4.11.Основнойкомпонентпчелиноговоска. Воска часто используются как защитный агент. Практически все зелёные части растений покрыты защитной восковой плёнкой. Прежде всего она защищает клетки от потери воды. Более толстый слой воска может защищать от механических повреждений. Если его ещё и выровнять, он будет выполнять роль светоотражающей поверхности. Это свойство используют многие фикусы для дополнительной защиты от избыточного солнечного света. Раньше такие листья использовались человеком в качестве бумаги. У растений спиртовая часть тоже остаточно длинная, до 40 атомов углерода. Но в состав возка входит обычно спектр таких сложных эфиров с разной длиной спиртовой части. Спермацет – особый воск, характерный для кашалотов. Он накапливается в больших количествах у них в голове и используется для регуляции глубины погружения. Этот воск содержит относительно короткие цепочки (С16-спирт и пальмитиновая кислота) и, соответственно, имеет более мягкую консистенцию и более низкую температуру плавления. При определённых температурах он изменяет свою плотность, изменяя и плавучесть кита. Если надо нырнуть на глубину, через спермацетовый орган пропускается холодная кровь, спермацет сжимается, голова становится более плотной и плавучесть понижается. Когда нужно всплыть, идёт противоположный процесс. Раньше спермацет использовался для изготовления различных мазей и кремов. Пока не появилось производство синтетических аналогов, кашалоты массово истреблялись. Иногда жирные кислоты присоединяются не сложноэфирными связями, а амидными связями к аминогруппе. NH2-группа даёт легко гидролизуемую связь, но из- за иного распределения электронной плотности такие соединения представлены одним соединением – сфингозином. Сфингозин содержит спиртовые и аминные группы, но при образовании сложных липидов он ведёт себя как амин. |