Липиды 200401 химия и обмен липидов
 Скачать 0.73 Mb. Скачать 0.73 Mb.
|

РОЛЬ УГЛЕВОДНЫХ КОМПОНЕНТОВ МЕМБРАНа) Участвуют в рецепции. б) Обеспечивают взаимодействие клеток друг с другом. в) Некоторые углеводные компоненты обеспечивают антигенную специфичность клеток. Например, эритроциты разных групп крови отличаются друг от друга по составу углеводных компонентов. Мембраны асимметричны. 2 монослоя отличаются друг от друга по своему составу. Например, гликолипиды плазматической мембраны всегда находятся в наружном монослое. Асимметрия характерна и для белковых компонентов. Аденилатциклаза. Ее активный центр находится на внутренней части мембраны. Белки-рецепторы свой углеводный компонент содержат с внешней стороны мембраны. Важнейшим компонентом плазматических мембран является холестерин. Холестерин взаимодействует с гидрофобными хвостами полярных молекул и ограничивает скорость диффузии липидов. Поэтому холестерин называют стабилизатором биологических мембран. Компоненты мембран не только движутся в пространстве, но и постоянно обновляются. Их место занимают новые молекулы. В учебную программу входит только обмен ГФЛ и холестерина. Липоиды синтезируются на мембранах эндоплазматического ретикулума. Наблюдается постоянное передвижение липоидов от мембран ЭПС к другим мембранам. СИНТЕЗ ХОЛЕСТЕРИНА Протекает в основном в печени на мембранах эндоплазматического ретикулума гепатоцитов. Этот холестерин - эндогенный. Происходит постоянный транспорт холестерина из печени в ткани. Для построения мембран используется также пищевой (экзогенный) холестерин. Ключевой фермент биосинтеза холестерина - ГМГ-редуктаза (бета-гидрокси, бета-метил, глутарил-КоА редуктаза). Этот фермент ингибируется по принципу отрицательной обратной связи конечным продуктом - холестерином. 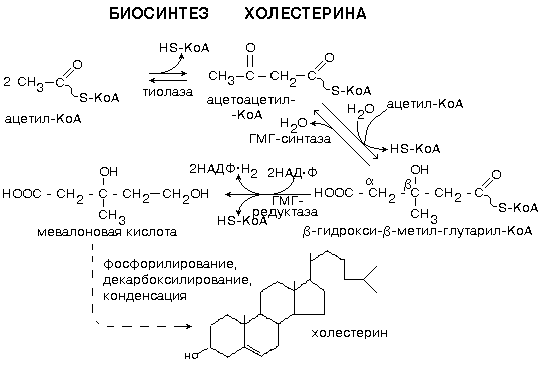 ТРАНСПОРТ ХОЛЕСТЕРИНА. Пищевой холестерин транспортируется хиломикронами и попадает в печень. Поэтому печень является для тканей источником и пищевого холестерина (попавшего туда в составе хиломикронов), и эндогенного холестерина. В печени синтезируются и затем попадают в кровь ЛОНП - липопротеины очень низкой плотности (состоят на 75% из холестерина), а также ЛНП - липопротеины низкой плотности(в их составе есть апобелок апоВ100. Почти во всех клетках имеются рецепторы для апоВ100. Поэтому ЛНП фиксируются на поверхности клеток. При этом наблюдается переход холестерина в клеточные мембраны. Поэтому ЛНП способны снабжать холестерином клетки тканей. Помимо этого, происходит и освобождение холестерина из тканей и транспорт его в печень. Транспортируют холестерин из тканей в печень липопротеины высокой плотности (ЛВП). Они содержат очень мало липидов и много белка. Синтез ЛВП протекает в печени. Частицы ЛВП имеют форму диска, и в их составе находятся апобелки апоА, апоС и апоЕ. В кровеносном русле к ЛНП присоединяется белок-фермент лецитинхолестеринацилтрансфераза (ЛХАТ) (смотрите рисунок). АпоС и апоЕ могут переходить от ЛВП на хиломикроны или ЛОНП. Поэтому ЛВП являются донорами апоЕ и апоС. АпоА является активатором ЛХАТ. ЛХАТ катализирует следующую реакцию: 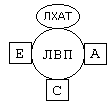 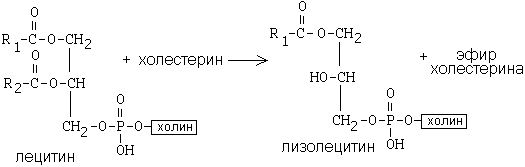 Это реакция переноса жирной кислоты из положения R2 на холестерин. Реакция является очень важной, потому что образующийся эфир холестерина является очень гидрофобным веществом и сразу переходит в ядро ЛВП - так при контакте с мембранами клеток ЛВП удаляют из них избыток холестерина. Дальше ЛВП идут в печень, там разрушаются, и избыток холестерина удаляется из организма. Нарушение соотношения между количеством ЛНП, ЛОНП и ЛВП может вызывать задержку холестерина в тканях. Это приводит к атеросклерозу. Поэтому ЛНП называют атерогенными липопротеинами, а ЛВП - антиатерогенными липопротеинами. При наследственном дефиците ЛВП наблюдаются ранние формы атеросклероза. ФУНКЦИИ ЛИПОИДОВ1. Структурная - составляют основу биологических мембран. 2. Регуляторная: а) вместе с белковыми компонентами обеспечивают избирательную проницаемость биологических мембран; б) при катаболизме липоидов образуются биологически активные вещества - регуляторы метаболизма. КАТАБОЛИЗМ ЛИПОИДОВ. Целью катаболизма является образование биологически активных веществ. Катаболизм холестерина происходит без разрушения полициклической структуры, в основном путем реакций микросомального окисления. Какие же биологически активные вещества образуются из холестерина? 1. Стероидные гормоны. 2. Желчные кислоты. 3. В коже под действием ультрафиолетового облучения образуется витамин D3. 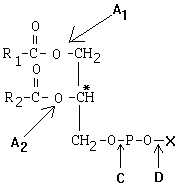 ГФЛ разрушаются путем гидролиза под действием фосфолипаз. Разные фосфолипазы обозначаются различными латинскими буквами. Какие биологически активные вещества образуются из ГФЛ? При образовании биологически активных веществ наибольшее значение имеют фосфолипазы "А2" и "С". Внутриклеточными посредниками при действии гормонов на клетку являются продукты гидролиза фосфолипазой "С" фосфорилированных фосфатидилинозитолов (диацилглицерин и инозитолфосфат) - смотрите лекцию "Гормоны". Фосфолипаза А2 отщепляет ненасыщенные жирные кислоты, которые могут быть предшественниками биологически активных веществ. Общим предшественником биологически активных веществ является АРАХИДОНОВАЯ КИСЛОТА, из которой образуется несколько групп этих веществ: ПГ (ПРОСТАГЛАНДИНЫ), Тх (ТРОМБОКСАНЫ), ЛТ (ЛЕЙКОТРИЕНЫ). Образуются эти вещества из полиненасыщенной арахидоновой кислоты в результате реакций ПЕРЕКИСНОГО ОКИСЛЕНИЯ. Впервые возможность перекисного окисления липидов (ПОЛ) была постулирована в 1887 году А.Н.Бахом. Арахидоновая кислота содержит 20 углеродных атомов и 4 двойные связи. В естественных условиях молекула арахидоновой кислоты имеет конфигурацию шпильки. 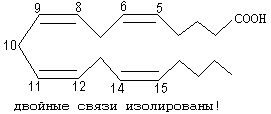 Арахидоновая кислота является субстратом для оксигеназ (ферменты, включающие кислород в состав субстрата). Перекисное окисление арахидоновой кислоты заключается в присоединении кислорода и образовании перекиси. Под действием активных форм кислорода происходит перемещение двойной связи и включение молекулы кислорода в молекулу арахидоновой кислоты. 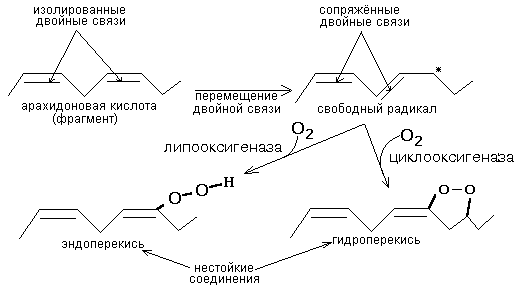 Перекиси - вещества нестойкие и быстро разрушаются. В липиде появляются "ОН"-группы или кетогруппы. В тканях человека и животных имеются два фермента перекисного окисления: ЦИКЛООКСИГЕНАЗА и ЛИПООКСИГЕНАЗА. При окислении с участием циклооксигеназы одновременно с окислением происходит циклизация, при действии липооксигеназы окисление идет без циклизации. Продукт действия циклооксигеназы: гидроперекись простагландин G2 (ПГG2). Затем он преобразуется в другую гидроперекись - ПГН2. Далее в результате действия других окислительных ферментов из ПГН2 образуются другие простагландины (они обозначаются латинскими буквами D, E, F и т.д.). Из ПГН2 образуется ещё 2 тромбоксана (Тх).  ПГ и Тх называются "ГОРМОНЫ МЕСТНОГО ДЕЙСТВИЯ". Регулирующий эффект оказывают там же, где образуются. В чем заключается их эффект? 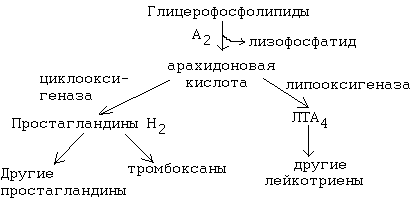 Простагландины влияют на проницаемость клеточных мембран. В частности, изменяют проницаемость кровеносных сосудов, влияют на тонус гладкомышечных клеток, на процессы реабсорбции в почках. Некоторые ПГ вызывают сокращение, а некоторые - расслабление мышечных элементов. Тромбоксаны регулируют функции тромбоцитов. Стимулируют агрегацию тромбоцитов. В результате действия липооксигеназы образуется лейкотриен А4 (ЛТА4). Это предшественник других лейкотриенов. Лейкотриены продуцируются лейкоцитами и регулируют функции лейкоцитов. В частности, вызывают хемотаксис гранулоцитов, стимулируют фагоцитоз. Способны влиять на клеточную проницаемость и на тонус гладких мышц. При различных аллергических заболеваниях (например, при бронхиальной астме) важную роль играет не только выделение гистамина, но и лейкотриенов. Лейкотриены обладают очень длительным воздействием на клетку. В животном организме может протекать и НЕФЕРМЕНТАТИВНОЕ ПЕРЕКИСНОЕ ОКИСЛЕНИЕ ЛИПИДОВ. Катализаторами являются в основном металлы с переменной валентностью (Fe, Cu, Ni). В отличие от ферментативного ПОЛ этот процесс может происходить с жирными кислотами, входящими в состав фосфолипидов, а не только со свободными ЖК. Включение кислорода в гидрофобные "хвосты" липидов приводит к появлению гидрофильности. Наступает набухание мембраны, нарушается ее проницаемость, и результатом может быть лизис клетки. В результате неферментативного ПОЛ может происходить разрушение гидрофобных "хвостов" липидов на более мелкие фрагменты. В результате образуется большое количество продуктов ПОЛ, многие из которых являются токсичными веществами. Усиление ПОЛ можно регистрировать по обнаружению этих низкомолекулярных продуктов. Один из показателей - повышение концентрации малонового диальдегида. Токсическое действие малонового альдегида заключается в его присоединении к аминогруппам белков и нуклеиновых кислот. В результате возможно нарушение биосинтеза и функционирования белков. |