Физиология. Механизмы регуляции работы сердца
 Скачать 0.61 Mb. Скачать 0.61 Mb.
|

|
В состоянии покоя сердце нагнетает от 4 до 6 л крови в минуту, за день -- до 8-10 тыс. л крови. Тяжёлая работа сопровождается 4-7-кратным увеличением перекачиваемого объёма крови. Показатели работы сердца рефлекторно изменяются в зависимости от напряжения О2и СО2в крови, от объема протекающей крови, от эмоционального состояния и физической нагрузки. Так, при физической нагрузке ударный объем может увеличиться в 2 - 3 раза, частота сокращений - в 3 - 4 раза, минутный объем кровообращения - в 4 - 5 раз. Механизмы регуляции работы сердца: 1. Интракардиальные: Внутриклеточные (геторометрический и гомеометрический механизмы) Межклеточные механизмы Интракардиальные сердечные рефлексы 2. Экстракардиальные: Нервные Гуморальные Интракардиальные механизмыв свою очередь подразделяются на миогенные (внутриклеточные), межклеточные и нервные (за счет внутрисердечной нервной системы). Внутриклеточные механизмы обусловлены свойствами кардиомиоцитов и лежат в основе закона Франка - Старлинга:чем больше приток крови, тем больше растягивается миокард во время диастолы, тем сильнее он сокращается во время систолы, т.е. чем больше крови поступает в желудочки, тем сильнее они потом сокращаются в систолу. Такой тип гемодинамической регуляции называется гетерометрическим. При растяжении напряжение развиваемое мышцей действительно увеличивается, но не за счет «увеличения зоны контакта актиновых и миозиновых протофибрил», а за счет увеличения вклада пассивного (эластичного) компонента в общее развиваемоемышечным волокном напряжение.Также этот механизм объясняется способностью Са2+ выходить из саркоплазматического ретикулума. Чем больше растянут саркомер, тем больше выделяется Са2+ в цитоплазму, обеспечивая большее сцепление актиновых и миозиновых нитей, и тем больше сила сокращений сердца. 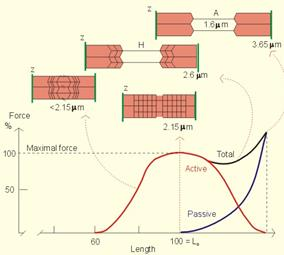 Рис. Зависимость между длиной саркомера, степенью перекрытия актиновых и миозиновых нитей и развитием напряжения для препарата одиночного волокна миоцита. Объяснение в тексте. Активное напряжение при растяжении саркомера более чем на 2,2 мкм уменьшается. Этот механизм служит для согласования систолических объёмов кровотока правой и левой половины сердца. Их систолический объём кровотока может изменяться от сокращения к сокращению. Если систолический объём левой половины во время какого-либо сокращения будет повышенным из-за значительного конечно-диастолического давления или объёма , при следующем сокращении ударный объём уменьшится и будет таким же, как и выброс правой половины сердца. Этот механизм саморегуляции включается при перемене положения тела, при резком увеличении объема циркулирующей крови (при переливании), а также при фармакологической блокад симпатической нервной системы бета-симпатолитиками. Гомеометрическая внутриклеточная регуляция сердца (феномен Анрепа и хронотропная зависимость Боудича) Гомеометрический механизм не зависит от исходной длины кардиомиоцитов. Сила сердечных сокращений может возрастать при увеличении частоты сокращений сердца. Чем чаще оно сокращается, тем выше амплитуда его сокращений («лестница» Боудича), однако при повышении ЧСС более 180 уд.мин сила сокращений снижается. Сердце человека и большинства животных, за исключением крыс в ответ на повышение ритма реагирует увеличением силы сокращений и, наоборот, с уменьшением ритма сила сокращений падает. Механизм этого феномена связан с накоплением или падением в миоплазме концентрации Са2 +, а следовательно - увеличением или уменьшением количества поперечных мостиков. При частом раздражении происходит увеличение ионов кальция в цитозоле, поскольку все больше ионов освобождаются из саркоплазматического ретикулума с каждым последующим мышечным потенциалом действия, а убрать их из саркоплазмы немедленно не удается ,т.к. это активный, а значит медленный процесс.  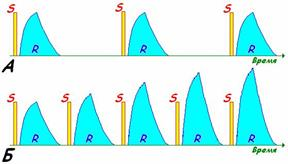 Рис. Возникновение «лестницы Боудича» при увеличении частоты следования импульсов. S - стимулы, одинаковые по силе, но разные по частоте (А - более редкие, Б - более частые). R - ответы (сокращения миокард) (А - одинаковые по амплитуде, Б - возрастающей амплитуды). При повышении давления в аорте до определенных пределов возрастает противонагрузка на сердце, происходит увеличение силы сердечных сокращений (феномен Анрепа), обеспечивая тем самым возможность выброса того же объема крови, что и при исходной величине артериального давления, т.е. чем больше противонагрузка, тем больше сила сокращений. Механизмы, лежащие в основе феномена Анрепа, до сих пор не раскрыты. Предполагают, что с увеличением противонагрузки растет концентрация Са2+ в межфибриллярном пространстве и поэтому возрастает сила сердечных сокращений. Регуляция межклеточных взаимодействий. Установлено, что вставочные диски, соединяющие клетки миокарда, имеют различную структуру. Одни участки вставочных дисков выполняют чисто механическую функцию, другие обеспечивают транспорт через мембрану кардиомиоцита необходимых ему веществ, третьи -- нексусы, или тесные контакты, проводят возбуждение с клетки на клетку. Нарушение межклеточных взаимодействий приводит к асинхронному возбуждению клеток миокарда и появлению сердечных аритмий. К межклеточным взаимодействиям следует отнести и взаимоотношения кардиомиоцитов с соединительнотканными клетками миокарда. Последние представляют собой не просто механическую опорную структуру. Они поставляют для сократительных клеток миокарда ряд сложных высокомолекулярных продуктов, необходимых для поддержания структуры и функции сократительных клеток. Подобный тип межклеточных взаимодействий получил название креаторных связей (Г. И. Косицкий). Внутрисердечные периферические рефлексы. Более высокий уровень внутриорганной регуляции деятельности сердца представлен внутрисердечными нервными механизмами. Обнаружено, что в сердце возникают так называемые периферические рефлексы, дуга которых замыкается не в ЦНС, а в интрамуральных ганглиях миокарда. После гомотрансплантации сердца теплокровных животных и дегенерации всех нервных элементов экстракардиального происхождения в сердце сохраняется и функционирует внутриорганная нервная система, организованная по рефлекторному принципу. Эта система включает афферентные нейроны, дендриты которых образуют рецепторы растяжения на волокнах миокарда и венечных (коронарных) сосудах, вставочные и эфферентные нейроны. Аксоны последних иннервируют миокард и гладкие мышцы коронарных сосудов. Указанные нейроны соединяются между собой синаптическими связями, образуя внутри-сердечные рефлекторные дуги. В экспериментах показано, что увеличение растяжения миокарда правого предсердия (в естественных условиях оно возникает при увеличении притока крови к сердцу) приводит к усилению сокращений миокарда левого желудочка. Таким образом, усиливаются сокращения не только того отдела сердца, миокард которого непосредственно растягивается притекающей кровью, но и других отделов, чтобы «освободить место» притекающей крови и ускорить выброс ее в артериальную систему. Доказано, что эти реакции осуществляются с помощью внутрисердечных периферических рефлексов (Г. И. Косицкий). Подобные реакции наблюдаются лишь на фоне низкого исходного кровенаполнения сердца и незначительной величины давления крови в устье аорты и коронарных сосудах. Если камеры сердца переполнены кровью и давление в устье аорты и коронарных сосудах высокое, то растяжение венозных приемников в сердце угнетает сократительную активность миокарда, в аорту выбрасывается меньшее количество крови, а приток крови из вен затрудняется. Подобные реакции играют важную роль в регуляции кровообращения, обеспечивая стабильность кровенаполнения артериальной системы. Гетерометрический и гомеометрический механизмы регуляции силы сокращения миокарда могут привести лишь к резкому увеличению энергии сердечного сокращения в случае внезапного повышения притока крови из вен или повышения артериального давления. Казалось бы, что при этом артериальная система не защищена от губительных для нее внезапных мощных ударов крови. В действительности же таких ударов не возникает благодаря защитной роли, осуществляемой рефлексами внутрисердечной нервной системы. Переполнение камер сердца притекающей кровью (равно как и значительное повышение давления крови в устье аорты, коронарных сосудов) вызывает снижение силы сокращений миокарда посредством внутрисердечных периферических рефлексов. Сердце при этом выбрасывает в артерии в момент систолы меньшее, чем в норме, количество содержащейся в желудочках крови. Задержка даже небольшого дополнительного объема крови в камерах сердца повышает диастолическое давление в его полостях, что вызывает снижение притока венозной крови к сердцу. Излишний объем крови, который при внезапном выбросе его в артерии мог бы вызвать пагубные последствия, задерживается в венозной системе. Опасность для организма представляло бы и уменьшение сердечного выброса, что могло бы вызвать критическое падение артериального давления. Такую опасность также предупреждают регуляторные реакции внутрисердечной системы. Недостаточное наполнение кровью камер сердца и коронарного русла вызывает усиление сокращений миокарда посредством внутрисердечных рефлексов. При этом желудочки в момент систолы выбрасывают в аорту большее, чем в норме, количество содержащейся в них крови. Это и предотвращает опасность недостаточного наполнения кровью артериальной системы. К моменту расслабления желудочки содержат меньшее, чем в норме, количество крови, что способствует усилению притока венозной крови к сердцу. В естественных условиях внутрисердечная нервная система не является автономной. Она -- лишь низшее звено сложной иерархии нервных механизмов, регулирующих деятельность сердца. Следующим, более высоким звеном этой иерархии являются сигналы, поступающие по блуждающим и симпатическим нервам, осуществляющие процессы экстракардиальной нервной регуляции сердца. Эффекты воздействий на сердце: влияние на ЧСС (т.е. на автоматизм) обозначают термином «хронотропноедействие» (может быть положительным и отрицательным), на силу сокращений (т.е. на сократимость) -- «инотропное действие» (положительное или отрицательное), на скорость предсердно-желудочкового проведения (что отражает функциюпроводимости) -- «дромотропное действие» (положительное или отрицательное), на возбудимость -- «батмотропное действие» (также положительное илиотрицательное). Внесердечные регуляторные механизмы Нервная экстракардиальная регуляция. Эта регуляция осуществляется импульсами, поступающими к сердцу из ЦНС по блуждающим и симпатическим нервам. Подобно всем вегетативным нервам, сердечные нервы образованы двумя нейронами. Тела первых нейронов, отростки которых составляют блуждающие нервы (парасимпатический отдел автономной нервной системы), расположены в продолговатом мозге. Отростки этих нейронов заканчиваются в интрамуральных ганглиях сердца. Здесь находятся вторые нейроны, отростки которых идут к проводящей системе, миокарду и коронарным сосудам. Первые нейроны симпатической части автономной нервной системы, передающие импульсы к сердцу, расположены в боковых рогах пяти верхних сегментов грудного отдела спинного мозга. Отростки этих нейронов заканчиваются в шейных и верхних грудных симпатических узлах. В этих узлах находятся вторые нейроны, отростки которых идут к сердцу. Большая часть симпатических нервных волокон, иннервирующих сердце, отходит от звездчатого узла. Парасимпатическая регуляция Влияние на сердце блуждающих нервов впервые изучили братья Вебер (1845). Они установили, что раздражение этих нервов тормозит работу сердца вплоть до полной его остановки в диастолу. Это был первый случай обнаружения в организме тормозящего влияния нервов. При электрическом раздражении периферического отрезка перерезанного блуждающего нерва происходит урежение сердечных сокращений. Это явление называется отрицательным хронотропным эффектом. Одновременно отмечается уменьшение амплитуды сокращений -- отрицательный инотропный эффект. При сильном раздражении блуждающих нервов работа сердца на некоторое время прекращается. В этот период возбудимость мышцы сердца понижена. Понижение возбудимости мышцы сердца называется отрицательным батмотропным эффектом. Замедление проведения возбуждения в сердце называется отрицательным дромотропным эффектом. Нередко наблюдается полная блокада проведения возбуждения в предсердно-желудочковом узле. Микроэлектродные отведения потенциалов от одиночных мышечных волокон предсердий показали увеличение мембранного потенциала -- гиперполяризацию при сильном раздражении блуждающего нерва При продолжительном раздражении блуждающего нерва прекратившиеся вначале сокращения сердца восстанавливаются, несмотря на продолжающееся раздражение. Это явление называют ускользанием сердца из-под влияния блуждающего нерва. Симпатическая регуляция Влияние на сердце симпатических нервов впервые было изучено братьями Цион (1867), а затем И. П. Павловым. Ционы описали учащение сердечной деятельности при раздражении симпатических нервов сердца (положительный хронотропный эффект); соответствующие волокна они назвали nn. accelerantes cordis (ускорители сердца). При раздражении симпатических нервов ускоряется спонтанная деполяризация клеток -- водителей ритма в диастолу, что ведет к учащению сердечных сокращений. Раздражение сердечных ветвей симпатического нерва улучшает проведение возбуждения в сердце (положительный дромотропный эффект) и повышает возбудимость сердца (положительный батмотропный эффект). Влияние раздражения симпатического нерва наблюдается после большого латентного периода (10 с и более) и продолжается еще долго после прекращения раздражения нерва. И. П. Павлов (1887) обнаружил нервные волокна (усиливающий нерв), усиливающие сердечные сокращения без заметного учащения ритма (положительный инотропный эффект). Инотропный эффект «усиливающего» нерва хорошо виден при регистрации внутрижелудочкового давления электроманометром. Выраженное влияние «усиливающего» нерва на сократимость миокарда проявляется особенно при нарушениях сократимости. Одной из таких крайних форм нарушения сократимости является альтернация сердечных сокращений, когда одно «нормальное» сокращение миокарда (в желудочке развивается давление, превышающее давление в аорте и осуществляется выброс крови из желудочка в аорту) чередуется со «слабым» сокращением миокарда, при котором давление в желудочке в систолу не достигает давления в аорте и выброса крови не происходит. «Усиливающий» нерв не только усиливает обычные сокращения желудочков, но и устраняет альтернацию, восстанавливая неэффективные сокращения до обычных. По мнению И. П. Павлова, эти волокна являются специально трофическими, т. е. стимулирующими процессы обмена веществ. Совокупность приведенных данных позволяет представить влияние нервной системы на ритм сердца как корригирующее, т. е. ритм сердца зарождается в его водителе ритма, а нервные влияния ускоряют или замедляют скорость спонтанной деполяризации клеток водителя ритма, ускоряя или замедляя таким образом частоту сердцебиений. В последние годы стали известны факты, свидетельствующие о возможности не только корригирующих, но и пусковых влияний нервной системы на ритм сердца, когда сигналы, приходящие по нервам, инициируют сокращения сердца. Это можно наблюдать в опытах с раздражением блуждающего нерва в режиме, близком к естественной импульсации в нем, т. е. «залпами» («пачками») импульсов, а не непрерывным потоком, как это делалось традиционно. При раздражении блуждающего нерва «залпами» импульсов сердце сокращается в ритме этих «залпов» (каждому «залпу» соответствует одно сокращение сердца). Меняя частоту и характеристику «залпов», можно управлять ритмом сердца в широких пределах. Воспроизведение сердцем центрального ритма резко изменяет электрофизиологические параметры деятельности синоатриального узла. При работе узла в режиме автоматии, а также при изменениях частоты под влиянием раздражения блуждающего нерва в традиционном режиме возбуждение возникает в одной точке узла, в случае воспроизведения центрального ритма в инициации возбуждения принимает участие одновременно множество клеток узла. На изохронной карте движения возбуждения в узле этот процесс отражается не в виде точки, а в виде большой площади, образованной одновременно возбуждающимися структурными элементами. Сигналы, обеспечивающие синхронное воспроизведение сердцем центрального ритма, отличаются по своей медиаторной природе от общетормозных влияний блуждающего нерва. По-видимому, выделяющиеся в этом случае наряду с ацетилхолином регуляторные пептиды отличаются по своему составу, т. е. реализация каждого типа эффектов блуждающего нерва обеспечивается своей смесью медиаторов («медиаторные коктейли»). 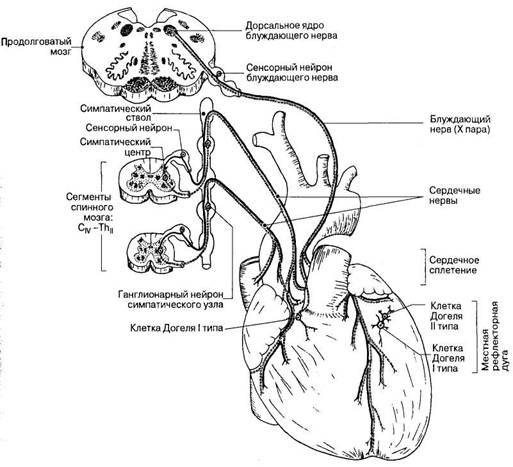 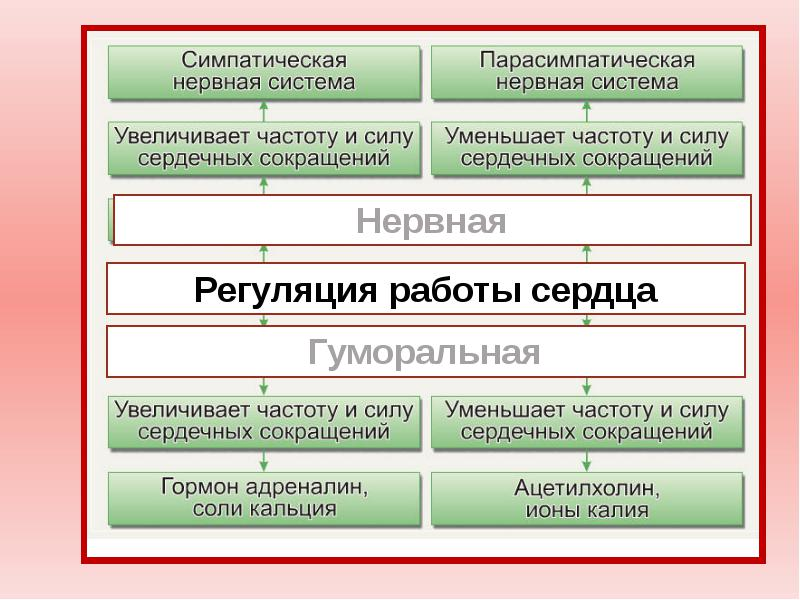 Гуморальная регуляция деятельности сердца Изменения работы сердца наблюдаются при действии на него ряда биологически активных веществ, циркулирующих в крови. Катехоламины (адреналин, норадреналин) увеличивают силу и учащают ритм сердечных сокращений, что имеет важное биологическое значение. При физических нагрузках или эмоциональном напряжении мозговой слой надпочечников выбрасывает в кровь большое количество адреналина, что приводит к усилению сердечной деятельности, крайне необходимому в данных условиях. Указанный эффект возникает в результате стимуляции катехоламинами рецепторов миокарда, вызывающей активацию внутриклеточного фермента аденилатциклазы, которая ускоряет образование 3',5'-циклического аденозинмонофосфата (цАМФ). Он активирует фосфорилазу, вызывающую расщепление внутримышечного гликогена и образование глюкозы (источника энергии для сокращающегося миокарда). Кроме того, фосфорилаза необходима для активации ионов Са2+ -- агента, реализующего сопряжение возбуждения и сокращения в миокарде (это также усиливает положительное инотропное действие катехоламинов). Помимо этого, катехоламины повышают проницаемость клеточных мембран для ионов Са2+, способствуя, с одной стороны, усилению поступления их из межклеточного пространства в клетку, а с другой -- мобилизации ионов Са2+ из внутриклеточных депо. Активация аденилатциклазы отмечается в миокарде и при действии глюкагона -- гормона, выделяемого α-клетками панкреатических островков, что также вызывает положительный инотропный эффект. Гормоны коры надпочечников, ангиотензин и серотонин также увеличивают силу сокращений миокарда, а тироксин учащает сердечный ритм. Гипоксемия, гиперкапния и ацидоз угнетают сократительную активность миокарда. Эндокринная функция сердца Миоциты предсердий образуют атриопептид, или натрийуретический гормон. Стимулируют секрецию этого гормона растяжение предсердий притекающим объемом крови, изменение уровня натрия в крови, содержание в крови вазопрессина, а также влияния экстракардиальных нервов. Натрийуретический гормон обладает широким спектром физиологической активности. Он сильно повышает экскрецию почками ионов Na+ и Сl-, подавляя их реабсорбцию в канальцах нефронов. Влияние на диурез осуществляется также за счет увеличения клубочковой фильтрации и подавления реабсорбции воды в канальцах. Натрийуретический гормон подавляет секрецию ренина, ингибирует эффекты ангиотензина II и альдостерона. Натрийуретический гормон расслабляет гладкие мышечные клетки мелких сосудов, способствуя тем самым снижению артериального давления, а также гладкую мускулатуру кишечника. Ситуационная задача 2.11 Выполните задачу с поэтапным решением. Ответы зафиксируйте в конспекте. В эксперименте установлено, что при раздражении усиливающего нерва Павлова наблюдается усиление сердечных сокращений. 1. Какой механизм регуляции появляется в этом эксперименте? 2. Какой отдел вегетативной нервной системы при этом активируется? 3.С действием какого медиатора связан указанный результат? 4. Какие клеточные механизмы при этом запускаются? Разбор лабораторной работы "Влияние растяжения волокон миокарда сердца лягушки на силу его сокращений (закон Старлинга)." Зависимость силы сердечных сокращений от степени растяжения клеток миокарда в период диастолы (закон Старлинга) традиционно демонстрировалась на изолированном сердце с помощью канюли Штрауба. Использование для этой цели полиэтиленовой канюли дает возможность выявить и четко записать указанную зависимость. Изменение исходного растяжения стенок желудочка на изолированном сердце регулируют с помощью смещения уровня жидкости в перфузионной канюле. Увеличение этого уровня сопровождается усилением сокращений желудочка и ростом амплитуды кардиограммы. Однако этот способ демонстрации закона сердца не лишен недостатков, которые очевидны при сопоставлении его с условиями эксперимента, в котором этот закон выявлен Старлингом, а именно: на сердечно-легочном препарате изменялся (увеличивался) венозный приток к правому сердцу при неизменном уровне сопротивления оттоку на выходе левого желудочка. В условиях изолированного сердца увеличивается не только сила растяжения желудочка в период диастолы, но также и сопротивление оттоку в систолу, то есть создаются условия для активации гомеометрического механизма регуляции. Поэтому целесообразно использовать для воспроизведения закона сердца на изолированном сердце лягушки импульсоподобное растяжение волокон миокарда в период диастолы с помощью пастеровской полиэтиленовой пипетки, заполненной раствором Рингера. Для работы необходимо: кимограф, штатив, операционный столик, препаровальный набор инструментов, лоток, рычажок Энгельмана, полиэтиленовая канюля, полиэтиленовая пипетка, раствор Рингера, лягушка. Ход работы. Обездвиживают лягушку, фиксируют на операционном столике брюшком кверху. Вскрытие грудной клетки и изоляцию сердца выполняют методом, описанным ранее. Канюлю с изолированным сердцем фиксируют с помощью резиновой трубки з верхней лапке штатива, а рычажок Энгельмана - на более низком уровне и соединяют с верхушкой сердца серфином и ниткой. Производится запись исходной кардиограммы на бумаге кимографа. Уровень жидкости в канюле устанавливают на расстоянии 2 см от начала шейки. Желательно сохранить его в течение всего опыта. Импуль-соподобное растяжение миокарда желудочка .моделируется в период диастолы с помощью полиэтиленовой пастеровской пипетки, заполненной раствором Рингера и опущенной в перфузионную канюлю до уровня шейки. Под визуальным контролем записи кардиограммы производят импульсоподобное давление на резиновый колпачок пастеровской пипетки в тот момент, когда рычажок Энгельмана совершает нисходящее колебание. Раствор Рингера под избыточным давлением переходит в полость желудочка и производит его растяжение — кривая кардиограммы начинает увеличиваться по амплитуде, а затем медленно возвращается к исходному уровню. Рекомендации к оформлению работы.Кратко запишите ход работы. Просмотрите виде экперимента. Объясните значение закона Старлинга для функции сердечно-сосудистой системы. |