шпаргалка по био. НУКЛЕИНОВЫЕ КИСЛОТЫ. Нуклеиновые кислоты. Структура и функции рнк и днк справочный материал нуклеиновые кислоты
 Скачать 197.52 Kb. Скачать 197.52 Kb.
|

1 2
НУКЛЕИНОВЫЕ КИСЛОТЫ. СТРУКТУРА И ФУНКЦИИ РНК И ДНК СПРАВОЧНЫЙ МАТЕРИАЛ НУКЛЕИНОВЫЕ КИСЛОТЫ Это гетерополимеры, мономерами которых служат нуклеотиды. Каждый нуклеотид состоит из остатка фосфорной кислоты, пятиуглеродпого сахара (рибозы или дезоксирибозы) и азотистого основания. Азотистые основания могут быть производными пурина (пуриновые основания - аденин и гуанин) или пиримидина (пиримидиновые основания - цитозин, тимин и урацил). В молекуле пуринов имеется два кольца, в молекуле пиримидинов - одно. В молекуле азотистого основания атомы углерода пронумерованы, в молекуле пентозы они обозначены цифрами с одним штрихом. В рибозе, в отличие от дезоксирибозы, с атомом 2’ связана ОН-группа. В результате присоединения азотистого основания к атому 1’ пентозы образуется нуклеозид. При присоединении к атому 5’ пентозы остатка фосфорной кислоты формируется нуклеотид. Адениловый нуклеотид содержит аденин, гуаниловый - гуанин, тимидиловый - тимин, цитидиловый - цитозин, уридиловый - урацил. Между пуриновыми и пиримидиновыми азотистыми основаниями могут образовываться водородные связи. Пары нуклеотидов с такими азотистыми основаниями называются комплементарными. Аденин комплементарен тимину и урацилу, гуанин - цитозину. Между гуанином и цитозином образуются три водородные связи, аденином и тимином или урацилом - две связи. Благодаря комплементарному взаимодействию азотистых оснований удерживаются вместе две нити ДНК и образуются петли в молекулах РНК. Нуклеотиды не только служат мономерами нуклеиновых кислот, но и входят в состав коферментов (НАД", НАДФ+, ФАД, ко-А, цАМФ), АТФ и сходных с ней производных нуклеотидов (ГТФ, ЦТФ). АТФ АТФ - аденозинтрифосфат. Молекула АТФ состоит из пуринового азотистого основания аденина, пятиуглеродного сахара рибозы и трех фосфатных групп. Богатую энергией связь между остатками фосфорной кислоты называют макроэргической. Под влиянием специфических ферментов АТФ может гидролизоваться с образованием АДФ и остатка фосфорной кислоты Фн. При этом выделяется около 30.6 кДж энергии на моль АТФ. Отщепление второй фосфатной группы дает столько же энергии, отщепление третьей - только 13.8 кДж/моль. Значение АТФ. АТФ - стандартная единица, в виде которой запасается энергия, высвобождаемая при окислении органических веществ. АТФ - постоянный источник энергии для клетки. Он мобилен и может доставлять химическую энергию в любую часть клетки. АТФ содержится во всех живых клетках и является универсальным носителем энергии. АТФ действует как связующее звено между процессами расщепления и синтеза. Из АДФ путем присоединения остатка фосфорной кислоты снова синтезируется АТФ, при этом, конечно, поглощается энергия ( 40 КДж/моль). РНК. РНК (рибонуклеиновая кислота) - это полимер, мономерами которого являются нуклеотиды. Нуклеотиды РНК состоят из пятиуглеродного сахара рибозы, остатка фосфорной кислоты и азотистого основания - цитозина, гуанина, аденина или урацила. Транспортную (т-РНК), состоящую из 80 - 100 нуклеотидов, содержащуюся в основном в цитоплазме клетки. Функция - перенос аминокислот к месту синтеза белка (в рибосомы). Полинуклеотидная цепь т-РНК образует три петли. т-РНК имеет два активных центра - один из них соединяется с определенной аминокислотой, другой находится на вершине средней петли, состоит из 3 нуклеотидов и служит для комплементарного соединения с молекулой и-РНК (антикодон). Рибосомную (р-РНК), состоящую из 3-5 тысяч нуклеотидов, составляющих основу рибосомы Информационную (и-РНК), содержащуюся в ядре и цитоплазме. Функция - перенос информации о структуре белка от ДНК к месту синтеза белка в рибосомах. Размер и-РНК зависит от длины участка ДНК, на котором они синтезированы (от 300 до 30 000 нуклеотидов). фермент эндонуклеаза "узнает" поврежденный участок ДНК; эндонуклеаза разрезает одну нить молекулы ДНК вблизи повреждения; фермепт окзонуклеаза вырезает поврежденный участок нити ДНК; фермент ДНК-полимераза проводит матричный синтез новой цепи (репаративную репликацию); фермент лигаза соединяет новообразованный участок с нитью ДНК. Пострепликативная репарация. Часть поврежденной ДНК не успевает репарироваться путем фотореактивации или эксцизии до репликации. В этом случае происходит пострепликативная репарация. При репликации поврежденный участок не может использоваться в качестве матрицы. Репликация вынуждена пропустить этот участок, оставляя брешь в дочерней нити. Таким образом, образующиеся дочерние молекулы ДНК различны по природе. Одна из них содержит неповрежденную родительскую цепь и нормальную дочернюю комплементарную цепь. Другая имеет родительскую цепь, несущую повреждение, и дочернюю цепь с брешью. Пострепликативная репарация происходит путем рекомбинации (обмена фрагментами) между дочерней, имеющей брешь, нитью и соответствующей ей нормальной материнской нитью другой молекулы. Образовавшийся в материнской цепи пробел может быть застроен обычным путем репаративного синтеза на комплементарной ей нормальной дочерней цепи. Поврежденный участок далее может быть исправлен путем фотореактивации или эксцизионной репарации. 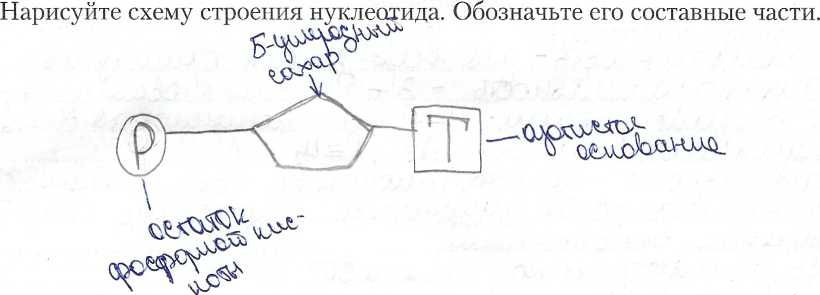 Задание 1. Задание 2. Нарисуйте схему строения молекулы АТФ. 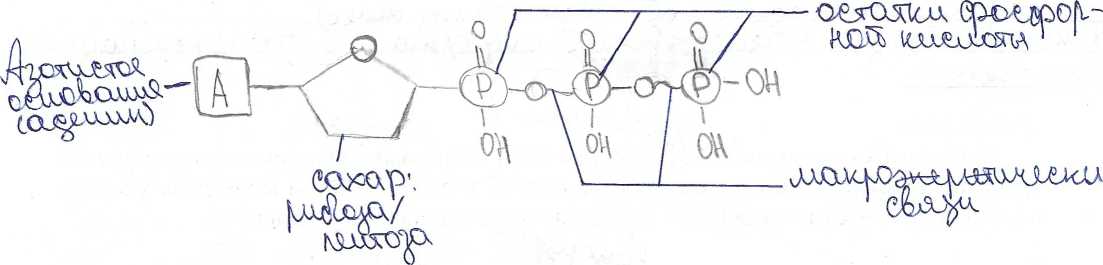 Задание 3. Задание 3.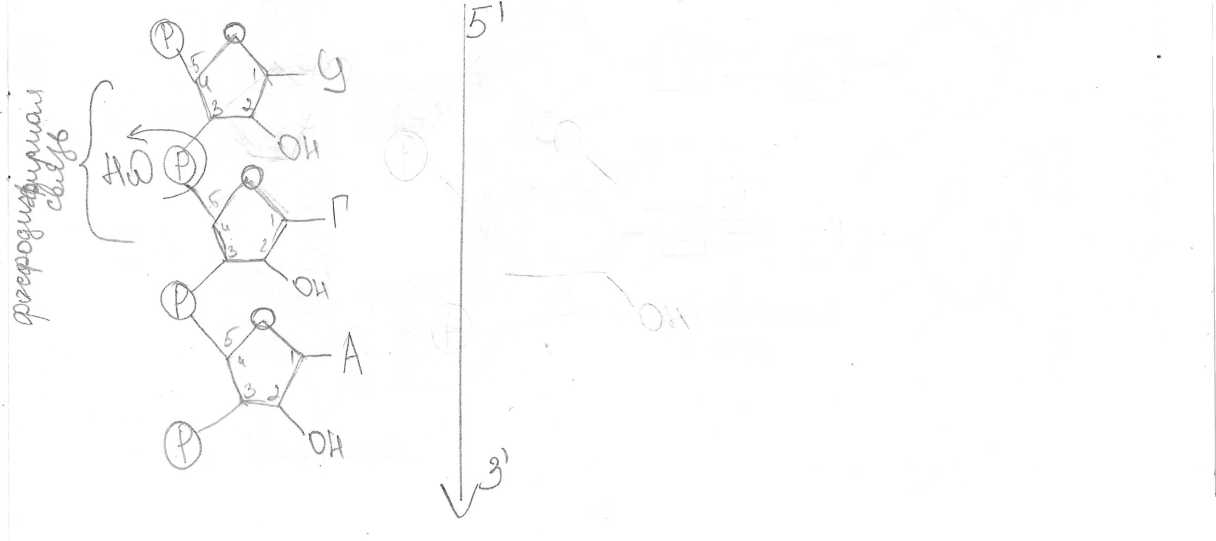 Нарисуйте схему первичной структуры РНК. Изобразите участок молекулы длиной три нуклеотида. На схеме обозначьте 3’ и 5’концы молекулы, укажите названия химических связей между основными компонентами молекулы. Нарисуйте схему первичной структуры РНК. Изобразите участок молекулы длиной три нуклеотида. На схеме обозначьте 3’ и 5’концы молекулы, укажите названия химических связей между основными компонентами молекулы.СПРАВОЧНЫЙ МАТЕРИАЛ Синтез белка осуществляется в два этапа: Транскрипция Трансляция Транскрипция - это синтез РНК на матрице ДНК. У эукариот происходит в ядре, у прокариот - в цитоплазме. В процессе транскрипции строится |^комплементарная копия одной из нитей ДНК (смысловой нити). В результате транскрипции синтезируются информационные (матричные), рибосомные и транспортные РНК. Транскрипцию осуществляет фермент РНК-полимераза. У эукариот транскрипцию осуществляют 3 разные РНК-полимеразы. , / РНК-полимераза I синтезирует рРНК, V РНК-полимераза II синтезирует иРНК, РНК-полимераза III синтезирует тРНК, 5SpPHK, малые ядерные РНК. РНК- полимераза связывается с молекулой ДНК в области промотора. Промотор - это участок ДНК, отмечающий начало транскрипции. Он расположен перед структурным геном. Присоединившись к промотору, РНК-полимераза раскручивает участок двойной спирали ДНК и разделяет комплементарные цепи. Одна из двух цепей - смысловая - служит матрицей для синтеза РНК. Нуклеотиды РНК комплементарны нуклеотидам смысловой цепи ДНК. Транскрипция идет от 5’-конца молекулы к ее З’-концу. РНК-полимераза отделяет синтезированный участок РНК от матрицы и восстанавливает двойную спираль ДНК. Тряцекри! 1лгист nprwin '[жяртгя до тех пор, пока РНК-полимераза не дойдет до терминатора. Терминатор - это участок ДНК, обозначающий конец транскрипции. Достигнув терминатора, РНК - полимераза отделяется и от матричной ДНК и от новосинтезированной молекулы РНК. Транскрипция делится на три этапа: инициацию, элонгацию и терминацию. 'Инициация - это присоединение РНК-полимеразы и помогающих ей белков- факторов транскрипции к ДНК и начало их работы. Элонгация - это наращивание полинуклеотидной цепи РНК. Терминация - окончание синтеза молекулы РНК. У прокариот новосинтезированные молекулы иРНК на 5’-конце имеют неинформативный участок, за которым следует инициирующий кодон АУГ, обозначающий начало трансляции. Далее находится информативный участок, на котором записана информация о структуре белка. За ним следует стоп-кодон (любой из трех: УАА, УАГ или УГА), определяющий конец трансляции. На 3’- коице находится концевая неинформативная последовательность. У эукариот новосинтезированная иРНК претерпевает посттранскрипционные изменения - процессинг. К 5’-концу пре-иРНК присоединяется кэп - метилированный гуанозинтрифосфат. К З’-концу присоединяется фрагмент, состоящий из 100-300 адениловых нуклеотидов - поли-А последовательность. Копирование (присоединение кэп) и полиаденилирование (присоединение поли-А последовательности) происходит в ядре в момент окончания синтеза иРНК. В результате образуется пре-иРНК, которая имеет следующее строение: на 5’-конце находится кэп, за ним неинформативная последовательность, инициирующий кодон АУГ, далее - отрезок, где информативные участки - экзоны - чередуются с неинформативными - нитронами. Этот отрезок заканчивается стоп-кодоном, за которым идет неинформативная последовательность и поли-А последовательность. У эукариот в ядре происходит сплайсинг - вырезание нитронов и сшивание экзонов. В результате процессинга формируется зрелая иРНК, которая через ядерные поры выходит из ядра в цитоплазму. В цитоплазме происходит трансляция. Трансляция - это процесс синтеза полипептидной цепи, проходящей на рибосоме. Трансляция происходит в цитоплазме. В эукариотических клетках рибосомы могут лежать в цитоплазме или могут прикрепляться к мембранам ЭПР. Рибосома состоит из двух субъединиц: большой и малой. Субъединицы построены из рРНК и белков. Прокариоты имеют рибосомы меньшего размера, чем эукариоты. Прокариотические рибосомы имеют константу седиментации 70S, эукариотические - 80S. Эукариоты в митохондриях и в пластидах имеют рибосомы 70S. Неактивная рибосома находится в цитоплазме в диссоциированном виде. Активная рибосома собирается из двух субъединиц, и при этом в ней образуются активные центры, в том числе - аминоацильный и пептидильный. В аминоацильном центре происходит взаимодействие кодона иРНК и антикодона тРНК. По антикодону узнается транспортная РНК, которая несет аминокислоту, зашифрованную кодоном. В пептидильном центре происходит образование пептидной связи. Транспортные РНК, доставляющие аминокислоты в рибосомы, всегда специфичны, т.е. одна тРНК может переносить только одну определенную аминокислоту. Это аминокислота зашифрована кодоном, которому комплементарен антикодон тРНК. Фермент аминоацил-тРНКсинтетаза активирует свободные аминокислоты и катализирует процесс присоединения аминокислот к соответствующим тРНК с образованием комплексов— аминоацил тРНК. В процессе трансляции рибосома переводит последовательность нуклеотидов иРНК в последовательность аминокислот полипептидной цепи. Трансляция дёлится на три этапа: инициация, элонгация и терминация. Инициация - это сборка рибосомы на инициирующем кодоне иРНК и начало ее работы. Инициация начинается с того, что с иРНК соединяется малая субъединица рибосомы и тРНК, несущая метионин, который соответствует инициирующему кодону АУГ. Затем к этому комплексу присоединяется большая субъединица. В результате инициирующий кодон оказывается^ пептидильном центре риоосомы, а в аминоацильном центре находится первый значащий кодон. К нему подходят различные тРНК, а останется в рибосоме только та тРНК, антикодон которой комплементарен кодону. Между комплементарными нуклеотидами кодона и антикодона образуются водородные связи. В итоге в рибосоме с иРНК оказываются временно связаны две тРНК. Каждая тРНК принесла в рибосому аминокислоту, зашифрованную кодоном иРНК. Между этими аминокислотами образуется пептидная связь. После этого тРНК, принесшая метионин, отделяется от своей аминокислоты и от иРНК и уходит из рибосомы. Рибосома перемещается на один триплет от 5’-конца к З’-концу иРНК. Элонгация - это процесс наращивания полипептидной цепочки. В аминоацильный центр рибосомы будут подходить различные тРНК. Процесс узнавания тРНК и процесс формирования пептидной связи будет повторятся до тех пор, пока в аминоациальном центре рибосомы не окажется стоп-кодон. Терминация - это завершение синтеза полипептида и диссоциация рибосомы на две субъединицы. Существует три стоп-кодона: УАА, УАГ и У ГА. Когда один из них оказывается в аминоацильном центре рибосомы, с ним связывается белок - фактор терминации трансляции. Это вызывает распад всего комплекса. СПРАВОЧНЫЙ МАТЕРИАЛ РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНОВ У ПРОКАРИОТ. ГИПОТЕЗА ОПЕРОНА I У прокариот известно несколько механизмов регуляции экспрессии генов: изменение дозы генов, регуляция на уровне транскрипции, посттрансляционная модификация полипептидов и т.д. Наиболее распространена регуляция на уровне транскрипции, которая по сравнению с другими способами регуляции очень экономна. В 1961 французские ученые Франсуа Жакоб, Жак Люсьен Моно и Андре Мишель Львов предложили гипотезу оперона, объясняющую регуляцию синтеза белков на уровне транскрипции у прокариот. Оперон - это совокупность тесно сцепленных структурных генов прокариот вместе с участком-оператором, регулирующим их транскрипцию. Структурные гены несут информацию о структуре белков, необходимых для выполнения одной функции. К оперону также относятся регуляторные участки: промотор и терминатор, которые участвуют в управлении транскрипцией. Промотор - это участок ДНК, определяющий начало транскрипции. С промотором связывается РНК-полимераза, которая затем продвигается вдоль ДНК, синтезируя молекулу РНК. Терминатор - это участок ДНК, обозначающий окончание транскрипции. РНК- полимераза, дойдя до терминатора, заканчивает транскрипцию и покидает ДНК. Оператор - это участок ДНК, с которым может специфически связываться белок- репрессор. Информацию о белке-репрессоре содержит ген-регулятор, который не входит в состав оперона. Связавшись с оператором, репрессор останавливает транскрипцию, т.к. препятствует продвижению РНК-полимеразы вдоль ДНК. Когда оперон активен, РНК-полимераза транскрибирует все структурные гены на одну молекулу иРНК. Информационная РНК, содержащая несколько генов, называется полицистронной. 1 2 |