Общие рекомендации
 Скачать 4.72 Mb. Скачать 4.72 Mb.
|

|
Процесс регенерации конечности у тритона/ саламандры. После ампутации регенерация конечности происходит строго упорядоченно, всегда одинаково. Восстанавливающийся конец округляется, затем приобретает коническую форму, растет в длину, становится похожим на ласт. Потом закладываются пальцы. К 8 неделе регенерация конечности полностью завершена. На клеточном уровне выделяют несколько фаз регенерации конечности:
Фаза заживления раны. В этот период происходит обрастание клетками раны на культе, возникает апикальная «шапочка» (если контакт нарушен – регенерации не будет). Процесс демонтирования. После заживления, в тканях, прилежащих к культе, происходит рассасывание ткани. Мышечные волокна утрачивают упорядоченность, становятся «растрепанными». В костной ткани утрачивается надкостница, появляются гигантские фагоцитирующие клетки, имеющие не менее 3-х ядер. Эти клетки захватывают матрикс и освобождают место для роста новой кости и хряща, удаляя ненужный материал. Концевая часть культи становится отечной и выпячивается. В культе накапливаются однотипные дедифференцированные клетки, уподобленные эмбриональным клеткам. Через некоторое время начинается деление дедифференцированных клеток. В отрастающую культю врастают нервы, и наступает стадия « конической бластемы». Конечность имеет форму ласта, нарастает клеточная масса, восстанавливается кровоток. Возникает «регенерационная почка». Фаза редифференцировки. Конечность удлиняется, начинается редифференцировка, и процесс регенерации подходит к концу. Если денервировать конечность - регенерация не произойдет т.к. нервная ткань выполняет эндокринную, проводящую функции. Кроме того, нервная ткань осуществляет секрецию белкового гормона, под контролем которого осуществляется регенерация. Репаративная (от лат. reparatio — восстановление) регенерация наступает после повреждения ткани или органа. Она очень разнообразна по факторам, вызывающим повреждения, по объемам повреждения, по способам восстановления. Механическая травма, например оперативное вмешательство, действие ядовитых веществ, ожоги, обморожения, лучевые воздействия, голодание, другие болезнетворные агенты,— все это повреждающие факторы. Наиболее широко изучена регенерация после механической травмы. Способность некоторых животных, таких, как гидра, планария, некоторые кольчатые черви, морские звезды, асцидия и др., восстанавливать утраченные органы и части организма издавна изумляла ученых. Ч. Дарвин, например, считал удивительными способность улитки воспроизводить голову и способность саламандры восстанавливать глаза, хвост и ноги именно в тех местах, где они отрезаны. Объем повреждения и последующее восстановление бывают весьма различными. Крайним вариантом является восстановление целого организма из отдельной малой его части, фактически из группы соматических клеток. Среди животных такое восстановление возможно у губок и кишечнополостных. Среди растений возможно развитие целого нового растения даже из одной соматической клетки, как это получено на примере моркови и табака. Такой вид восстановительных процессов сопровождается возникновением новой морфогенетической оси организма и назван Б.П. Токиным «соматическим эмбриогенезом», ибо во многом напоминает эмбриональное развитие. Существуют примеры восстановления больших участков организма, состоящих из комплекса органов. В качестве примера служат регенерация ротового конца у гидры, головного конца у кольчатого червя и восстановление морской звезды из одного луча (рис. 8.24). Широко распространена регенерация отдельных органов, например конечности у тритона, хвоста у ящерицы, глаз у членистоногих. Заживление кожных покровов, ран, повреждений костей и других внутренних органов является менее объемным процессом, но не менее важным для восстановления структурно-функциональной целостности организма. Особый интерес представляет способность зародышей на ранних стадиях развития восстанавливаться после значительной утраты материала. Эта способность была последним аргументом в борьбе между сторонниками преформизма и эпигенеза и привела в 1908 г. Г. Дриша к концепции эмбриональной регуляции. 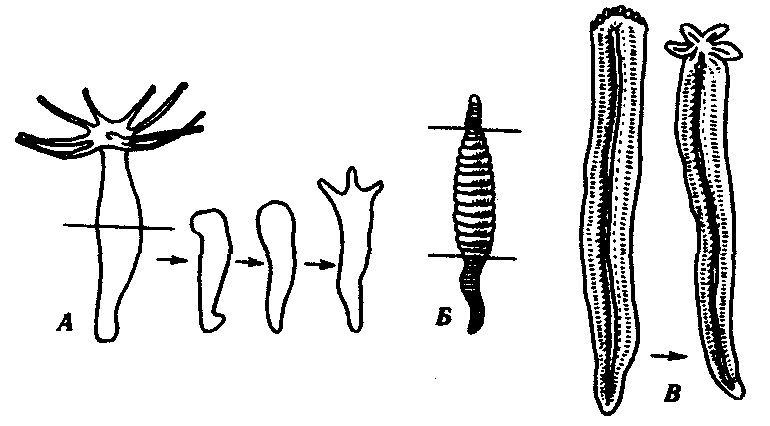 Рис. 8.24. Регенерация комплекса органов у некоторых видов беспозвоночных животных. А — гидра;Б — кольчатый червь; В — морская звезда (пояснение см. в тексте) Существует несколько разновидностей или способов репаративной регенерации. К ним относят эпиморфоз, морфаллаксис, заживление эпителиальных ран, регенерационную гипертрофию, компенсаторную гипертрофию. Эпителизация при заживлении ран с нарушенным эпителиальным покровом идет примерно одинаково, независимо от того, будет далее происходить регенерация органа путем эпиморфоза или нет. Эпидермальное заживление раны у млекопитающих в том случае, когда раневая поверхность высыхает с образованием корки, проходит следующим образом (рис. 8.25). Эпителий на краю раны утолщается вследствие увеличения объема клеток и расширения межклеточных пространств. Сгусток фибрина играет роль субстрата для миграции эпидермиса в глубь раны. В мигрирующих эпителиальных клетках нет митозов, однако они обладают фагоцитарной активностью. Клетки с противоположных краев вступают в контакт. Затем наступает кератинизация раневого эпидермиса и отделение корки, покрывающей рану. 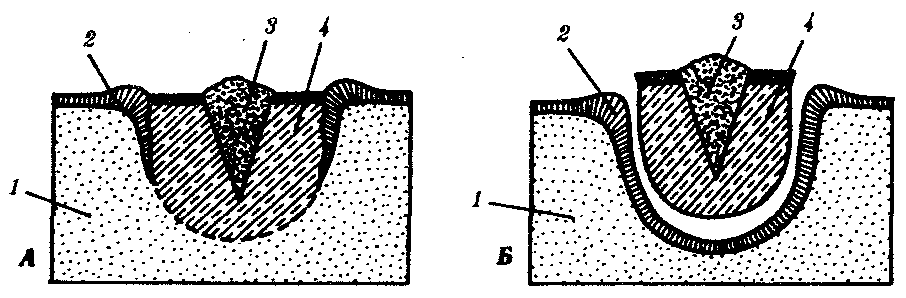 Рис. 8.25. Схема некоторых событий, происходящих при эпителизации кожной раны у млекопитающих. А—начало врастания эпидермиса под некротическую ткань; Б— срастание эпидермиса и отделение струпа: 1—соединительная ткань, 2—эпидермис, 3—струп, 4—некротическая ткань К моменту встречи эпидермиса противоположных краев в клетках, расположенных непосредственно вокруг края раны, наблюдается вспышка митозов, которая затем постепенно падает. По одной из версий, эта вспышка вызвана понижением концентрации ингибитора митозов — кейлона. Эпиморфоз представляет собой наиболее очевидный способ регенерации, заключающийся в отрастании нового органа от ампутационной поверхности. Регенерация конечности тритона и аксолотля изучена детально. Выделяют регрессивную и прогрессивную фазы регенерации. Регрессивная фаза начинается с заживления раны, во время которого происходят следующие основные события: остановка кровотечения, сокращение мягких тканей культи конечности, образование над раневой поверхностью сгустка фибрина и миграция эпидермиса, покрывающего ампутационную поверхность. Затем начинается разрушение остеоцитов на дистальном конце кости и других клеток. Одновременно в разрушенные мягкие ткани проникают клетки, участвующие в воспалительном процессе, наблюдается фагоцитоз и местный отек. Затем вместо образования плотного сплетения волокон соединительной ткани, как это происходит при заживлении ран у млекопитающих, в области под раневым эпидермисом утрачиваются дифференцированные ткани. Характерна остеокластическая эрозия кости, что является гистологическим признаком дедифференцировки. Раневой эпидермис, уже пронизанный регенерирующими нервными волокнами, начинает быстро утолщаться. Промежутки между тканями все более заполняются мезенхимоподобными клетками. Скопление мезенхимных клеток под раневым эпидермисом является главным показателем формирования регенерационной бластемы. Клетки бластемы выглядят одинаково, но именно в этот момент закладываются основные черты регенерирующей конечности. Затем начинается прогрессивная фаза, для которой наиболее характерны процессы роста и морфогенеза. Длина и масса регенерационной бластемы быстро увеличиваются. Рост бластемы происходит на фоне идущего полным ходом формирования черт конечности, т.е. ее морфогенеза. Когда форма конечности в общих чертах уже сложилась, регенерат все еще меньше нормальной конечности. Чем крупнее животное, тем больше эта разница в размерах. Для завершения морфогенеза требуется время, по истечении которого регенерат достигает размеров нормальной конечности. Некоторые стадии регенерации передней конечности у тритона после ампутации на уровне плеча показаны на рис. 8.26. Время, необходимое для полной регенерации конечности, варьирует в зависимости от размера и возраста животного, а также от температуры, при которой она протекает. 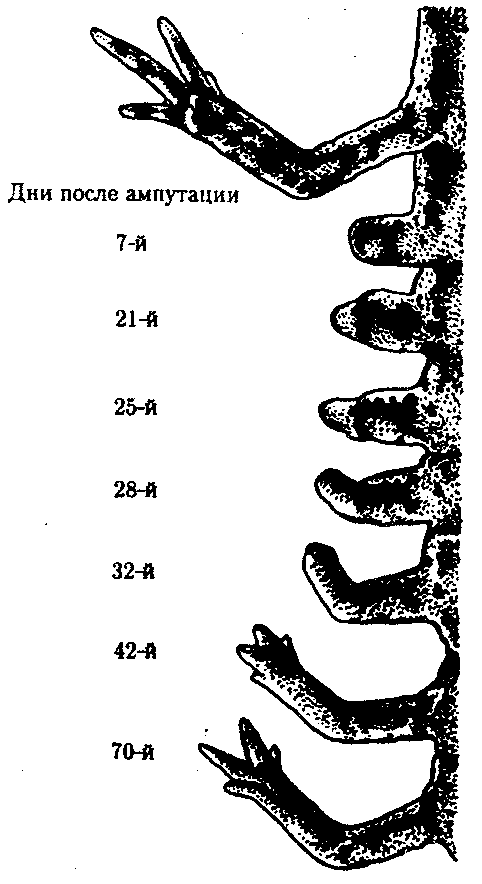 Рис. 8.26. Стадии регенерации передней конечности у тритона У молодых личинок аксолотлей конечность может регенерировать за 3 нед, у взрослых тритонов и аксолотлей за 1—2 мес, а у наземных амбистом для этого требуется около 1 года. При эпиморфной регенерации не всегда образуется точная копия удаленной структуры. Такую регенерацию называют атипичной. Существует много разновидностей атипичной регенерации. Гипоморфоз — регенерация с частичным замещением ампутированной структуры. Так, у взрослой шпорцевой лягушки возникает шиловидная структура вместо конечности. Гетероморфоз — появление иной структуры на месте утраченной. Это может проявляться в виде гомеозисной регенерации, заключающейся в появлении конечности на месте антенн или глаза у членистоногих, а также в изменении полярности структуры. Из короткого фрагмента планарии можно стабильно получать биполярную планарию (рис. 8.27). Встречается образование дополнительных структур, или избыточная регенерация. После надреза культи при ампутации головного отдела планарии возникает регенерация двух голов или более (рис. 8.28). Можно получить больше пальцев при регенерации конечности аксолотля, повернув конец культи конечности на 180°. Дополнительные структуры являются зеркальным отражением исходных или регенерировавших структур, рядом с которыми они расположены (закон Бэйтсона). 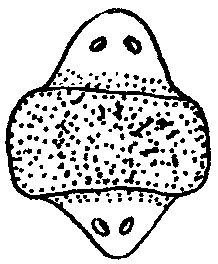 Рис. 8.27. Биполярная планария Морфаллаксис — это регенерация путем перестройки регенерирующего участка. Примером служит регенерация гидры из кольца, вырезанного из середины ее тела, или восстановление планарии из одной десятой или двадцатой ее части. На раневой поверхности в этом случае не происходит значительных формообразовательных процессов. Отрезанный кусочек сжимается, клетки внутри него перестраиваются, и возникает целая особь уменьшенных размеров, которая затем растет. Этот способ регенерации впервые описал Т. Морган в 1900 г. В соответствии с его описанием морфаллаксис осуществляется без митозов. Нередко имеет место сочетание эпиморфного роста на месте ампутации с реорганизацией путем морфаллаксиса в прилежащих частях тела.  Рис. 8.28. Многоголовая планария, полученная после ампутации головы и нанесения насечек на культю Регенерационная гипертрофия относится к внутренним органам. Этот способ регенерации заключается в увеличении размеров остатка органа без восстановления исходной формы. Иллюстрацией служит регенерация печени позвоночных, в том числе млекопитающих. При краевом ранении печени удаленная часть органа никогда не восстанавливается. Раневая поверхность заживает. В то же время внутри оставшейся части усиливается размножение клеток (гиперплазия) и в течение двух недель после удаления 2/3 печени восстанавливаются исходные масса и объем, но не форма. Внутренняя структура печени оказывается нормальной, дольки имеют типичную для них величину. Функция печени также возвращается к норме. Компенсаторная гипертрофия заключается в изменениях в одном из органов при нарушении в другом, относящемся к той же системе органов. Примером является гипертрофия в одной из почек при удалении другой или увеличение лимфатических узлов при удалении селезенки. Последние два способа отличаются местом регенерации, но механизмы их одинаковы: гиперплазия и гипертрофия. Восстановление отдельных мезодермальных тканей, таких, как мышечная и скелетная, называют тканевой регенерацией. Для регенерации мышцы важно сохранение хотя бы небольших ее культей на обоих концах, а для регенерации кости необходима надкостница. Регенерация путем индукции происходит в определенных мезодермальных тканях млекопитающих в ответ на действие специфических индукторов, которые вводят внутрь поврежденной области. Этим способом удается получить полное замещение дефекта костей черепа после введения в него костных опилок. Таким образом, существует множество различных способов или типов морфогенетических явлений при восстановлении утраченных и поврежденных частей организма. Различия между ними не всегда очевидны, и требуется более глубокое понимание этих процессов. Изучение регенерационных явлений касается не только внешних проявлений. Существует целый ряд вопросов, носящих проблемный и теоретический характер. К ним относятся вопросы регуляции и условий, в которых протекают восстановительные процессы, вопросы происхождения клеток, участвующих в регенерации, способности к регенерации у различных групп, животных и особенностей восстановительных процессов у млекопитающих. Установлено, что в конечности амфибий после ампутации и в процессе регенерации происходят реальные изменения электрической активности. При проведении электрического тока через ампутированную конечность у взрослых шпорцевых лягушек наблюдается усиление регенерации передних конечностей. В регенератах увеличивается количество нервной ткани, из чего делается вывод, что электрический ток стимулирует врастание нервов в края конечностей, в норме не регенерирующих. Попытки стимулировать подобным образом регенерацию конечностей у млекопитающих оказались безуспешными. Так, под действием электрического тока или при сочетании действия электрического тока с фактором роста нервов удавалось получить у крысы только разрастание скелетной ткани в виде хрящевых и костных мозолей, которые не походили на нормальные элементы скелета конечностей. Несомненна регуляция регенерационных процессов со стороны нервной системы. При тщательной денервации конечности во время ампутации эпиморфная регенерация полностью подавляется и бластема никогда не образуется. Были проведены интересные опыты. Если нерв конечности тритона отвести под кожу основания конечности, то образуется дополнительная конечность. Если его отвести к основанию хвоста — стимулируется образование дополнительного хвоста. Отведение нерва на боковую область никаких дополнительных структур не вызывает. Эти эксперименты привели к созданию концепции регенерационных полей. . Было установлено, что для инициации регенерации решающим является число нервных волокон. Тип нерва роли не играет. Влияние нервов на регенерацию связывается с трофическим действием нервов на ткани конечностей. Получены данные в пользу гуморальной регуляции регенерационных процессов. Особенно распространенной моделью для изучения этого является регенерирующая печень. После введения нормальным интактным животным сыворотки или плазмы крови от животных, подвергшихся удалению печени, у первых наблюдалась стимуляция митотической активности клеток печени. Напротив, при введении травмированным животным сыворотки от здоровых животных получали снижение количества митозов в поврежденной печени. Эти опыты могут свидетельствовать как о присутствии в крови травмированных животных стимуляторов регенерации, так и о присутствии в крови интактных животных ингибиторов клеточного деления. Объяснение результатов опытов затрудняется необходимостью учитывать иммунологический эффект инъекций. Важнейшим компонентом гуморальной регуляции компенсаторной и регенерационной гипертрофии является иммунологический ответ. Не только частичное удаление органа, но и многие воздействия вызывают возмущения в иммунном статусе организма, появление аутоантител и стимуляцию процессов клеточной пролиферации. Большие разногласия существуют по вопросу о клеточных источниках регенерации. Откуда берутся или как возникают недифференцированные клетки бластемы, морфологически сходные с мезенхимными? Существует три предположения. 1. Гипотеза резервных клеток подразумевает, что предшественниками регенерационной бластемы являются так называемые резервные клетки, которые останавливаются на некоем раннем этапе своей дифференцировки и не участвуют в процессе развития до получения стимула к регенерации. 2. Гипотеза временной дедифференцировки, или модуляции, клеток предполагает, что в ответ на регенерационный стимул дифференцированные клетки могут утрачивать признаки специализации, но затем снова дифференцируются в тот же клеточный тип, т.е., потеряв на время специализацию, они не утрачивают детерминацию. 3. Гипотеза полной дедифференцировки специализированных клеток до состояния, сходного с мезенхимными клетками и с возможной последующей трансдифференцировкой или метаплазией, т.е. превращением в клетки другого типа, полагает, что в этом случае клетка утрачивает не только специализацию, но и детерминацию. Современные методы исследования не позволяют с абсолютной достоверностью доказать все три предположения. Тем не менее абсолютно верно, что в культях пальцев аксолотля происходит высвобождение хондроцитов из окружающего матрикса и миграция их в регенерационную бластему. Дальнейшая их судьба не определена. Большинство исследователей признают дедифференцировку и метаплазию при регенерации хрусталика у амфибий. Теоретическое значение этой проблемы заключается в допущении возможности или невозможности изменений клеткой ее программы до такой степени, что она возвращается в состояние, когда снова способна делиться и репрограммироватьсвой синтетический аппарат. Например, хондроцит становится миоцитом или наоборот. Способность к регенерации не имеет однозначной зависимости от уровня организации, хотя давно уже было замечено, что более низко организованные животные обладают лучшей способностью к регенерации наружных органов. Это подтверждается удивительными примерами регенерации гидры, планарий, кольчатых червей, членистоногих, иглокожих, низших хордовых, например асцидий. Из позвоночных наилучшей регенерационной способностью обладают хвостатые земноводные. Известно, что разные виды одного и того же класса могут сильно отличаться по способности к регенерации. Кроме того, при изучении способности к регенерации внутренних органов оказалось, что она значительно выше у теплокровных животных, например у млекопитающих, по сравнению с земноводными. Регенерация у млекопитающих отличается своеобразием. Для регенерации некоторых наружных органов нужны особые условия. Язык, ухо, например, не регенерируют при краевом повреждении. Если же нанести сквозной дефект через всю толщу органа, восстановление идет хорошо. В некоторых случаях наблюдали регенерацию сосков даже при ампутации их по основанию. Регенерация внутренних органов может идти очень активно. Из небольшого фрагмента яичника восстанавливается целый орган. Об особенностях регенерации печени уже было сказано выше. Различные ткани млекопитающих тоже хорошо регенерируют. Есть предположение, что невозможность регенерации конечностей и других наружных органов у млекопитающих носит приспособительный характер и обусловлена отбором, поскольку при активном образе жизни нежные морфогенетические процессы затрудняли бы существование. Достижения биологии в области регенерации успешно применяются в медицине. Однако в проблеме регенерации очень много нерешенных вопросов. 75.Биологическое и медицинское значение проблемы регенерации. Проявление регенерационной способности у человека. Регенерация патологически измененных органов и обратимость патологически изменённых органов. Регенерационная терапия. При разрезе в рану устремляется кровь, лейкоциты которой запускают воспалительный процесс. Клетки прилежащей эпителиальной ткани делятся и образуют «струп» (рубец). Потом начинается процесс заживления. В настоящее время интенсивно изучаются проблемы регенерации, особенно связанные с медициной. Стволовые клетки обладают свойствами: - стволовая клетка не является окончательно дифференцированной (она скорее детерминирована); - стволовая клетка способна к неограниченному делению; - при делении часть клеток остается стволовыми, другая часть подвергается процессу дифференцировки. Центров по применению стволовых клеток очень мало, в России существует только 2 таких центра. Однако стволовые клетки есть везде. Для лечения и экспериментов берется пуповинная кровь с целью получения стволовых клеток. Кости черепа в норме не регенерируют. Под руководством И.И.Полежаева происходило удаление участка 10х10 см черепа собаки. Из кости получали путем измельчения костные опилки, которые помещали на рану. В другом эксперименте использовали костные опилки донора и кровь реципиента. Через неделю происходило рассасывание опилок, а к концу 1 года рана зарастала. Большое значение имеет регенерация после радиоактивного облучения. Малые дозы стимулируют, а большие, наоборот ингибируют данный процесс. Если провести механическое раздавливание культи или помещение ее в кислоту – регенерация идет в 50% случаев. Елизаров проводил ломку и удлинение костей. Им были созданы уникальные аппараты, благодаря которым было возможно раздвижение костей скелета и коррекция их формы. Остро стоит проблема регенерации печени. При циррозе печени приходится проводить ее частичное удаление. Иногда подобная операция проводится несколько раз, печень быстро регенерирует без сохранения формы, сохраняя функцию и общую массу. Регенерацию можно стимулировать антикейлоном, витамином В12, АТФ, РНК. Выделяют типы регенерации в патологически измененных органах.
76.Понятие о гомеостазе. Общие закономерности гомеостаза живых систем. Генетические, клеточные и системные основы гомеостатических реакций организма. Роль эндокринной, нервной и иммунной систем в обеспечении гомеостаза и адаптивных изменений. Термин «гомеостаз» был предложен для понимания постоянства состава лимфы, крови и тканевой жидкости. Гомеостаз характерен для любой системы, это своего рода обобщение множества частных проявлений стабильности системы. Как же организм сохраняет постоянство?? Гомеостаз – поддержание постоянства внутренней среды организма в непрерывно изменяющихся условиях внешней среды. Т.к. организм – многоуровневый саморегулирующийся объект, его можно рассматривать с точки зрения кибернетики. Тогда, организм – сложная многоуровневая саморегулирующаяся система с множеством переменных. Переменные входа: - причина; - стимул; - раздражение. Переменные выхода: - эффект; - ответ; - реакция; - следствие. Причина – отклонение от нормы реакции в организме. Решающая роль принадлежит обратной связи. Существует положительная и отрицательная обратная связь. Отрицательная обратная связь уменьшает действие входного сигнала на выходной. Положительная обратная связь увеличивает действие входного сигнала на выходной эффект действия. Живой организм – ультрастабильная система, осуществляющая поиск наиболее оптимального устойчивого состояния, которое обеспечивается адаптациями. Адаптация – поддержание переменных показателей на поведенческом, анатомическом, биохимическом и других уровнях. Этология – наука, изучающая поведение животных и человека. Типы поведения животных и человека ограничены их морфологическими и физиологическими особенностями. У человека есть зависимость поведения от типа сложения. Существуют 3 типа сложения:
Животные могут совершенствовать свои движения за счет информации, кроме того, они имеют возможность регулировать их. Животные должны различать объекты внешней среды, получать информацию при помощи органов чувств. Полученная информация подвергается переработке нервной и эндокринной системами. Многие типы поведения могут вызывать гормональные изменения. Морфологические и физиологические признаки подвержены естественному отбору, поведение в свою очередь, зависит от этих признаков, а значит, зависит и от естественного отбора. Поведение передается по наследству, повышает приспособляемость, увеличивает продолжительность жизни, количество потомков. Различные поведенческие реакции позволяют использовать благоприятные условия среды, защищают организм от неблагоприятных условий. Например, у пчел поддержание чистоты в улье. За гигиеническое поведение отвечают как минимум 2 гена. Поддержание чистоты защищает пчел от болезней. Поведение ящерицы, отбрасывающей хвост, если это необходимо, - тоже приспособительная реакция. Другие типы поведенческих реакций наблюдаются при защите от хищников, при поиске пищи, партнера, защите потомства и многих других случаях. Некоторые насекомые выделяют особые химические вещества – феромоны для привлечения потомства. В брачный период лягушки квакают и их «песня» видоспецифична. Поведенческие признаки обладают не только адаптивными свойствами, но могут также и наследоваться, что обусловливает естественный отбор. Не все типы поведения получаются при передаче с генами, они могут приобретаться – благоприобретенные. Резкую границу между теми и другими провести нельзя, т.к. гены и среда тесно взаимодействуют друг с другом, поэтому выделить отдельно генетические и благоприобретенные свойства нельзя. Можно привести следующие примеры генетических свойств. Хорея Гентингтона – наследственное заболевание, «танец», поражает ЦНС, у больных также нарушена пространственная ориентация. Другой пример, дауны бывают благожелательными, ласковыми, подражают действиям здоровых людей Итак, важные свойства поведенческих реакций:
Если организм не смог адаптироваться на поведенческом уровне, он делает это на биохимическом уровне. Биохимическая адаптация очень сложна, наиболее характерна для растений, т.к. животному проще мигрировать. Процесс адаптации бывает по времени: - эволюционная адаптация; - акклиматизация; - немедленная адаптация. Эволюционная адаптация – длительный процесс, приобретение новой генетической информации, изменяется генотип, следовательно, изменяется и фенотип. Для своего завершения подобная адаптация требует многих поколений. Акклиматизация – адаптации, которые происходят в процессе жизни в естественных условиях. Акклимация – адаптации, происходящие в искусственных условиях. Происходит в течение нескольких часов – лет (зима – лето). Смена часовых поясов, перевод времени. Немедленная адаптация сопровождается почти мгновенной адаптивной реакцией (психогенное воздействие, переход из тепла в холод). Кратковременная реакция. Любая адаптация возникает в результате взаимодействия генетических факторов и факторов внешней среды. Генетический аспект гомеостаза рассматривают с 3 позиций: - гомеостаз генотипа; - гомеостаз организма как целого. Контроль за единством генотипа всего организма. Поддержание гомеостаза осуществляется при гибели видоизмененных клеток. - гомеостаз популяции. Закон генетической стабильности в популяции. В поддержании гомеостаза участвуют различные системы. Нервная сигнализация – основной инструмент передачи и оценки сигналов из внутренней и внешней среды. Гормоны принимают участие в регуляции гомеостаза. Регулируют обмен веществ, воды, белков, липидов, углеводов, энергии, электролитов. Контролируют работу всех органов, в том числе почек, печени, ЦНС. Иммунная система защищает постоянство внутренней среды организма от факторов 2-х групп: - микроорганизмов и экзогенных факторов с признаками чужеродной генетической информацией; - соматических мутаций. Достаточно изменений в 1-2 генах, чтобы сработала иммунная система. 77.Проблемы трансплантации органов и тканей. Ауто-, алло- и ксенотрансплантация, трансплантация жизненно важных органов. Иммунитет. Тканевая несовместимость и пути ее преодоления. Искусственные органы. Из-за бурного развития трансплантологии остро встал вопрос о трансплантационном иммунитете. Трансплантология – медико-биологическая наука, изучающая вопросы заготовки, консервирования и пересадки органов и тканей. Трансплантационный иммунитет – своеобразная реакция организма на трансплантацию, проявляющаяся в отторжении пересаженных органов и тканей. Классификация терминов (Вена, 1967 год). Трансплантат – пересаживаемая ткань или орган. Реципиент – тот, кому пересаживается орган или ткань. Донор – тот, от кого берут трансплантат. Аутотрансплантация – пересадка тканей и органов в пределах одного организма (в таком случае говорят об аутотрансплантате) Изотрансплантация (изотрансплантат) - пересадка тканей и органов между организмами, идентичными по генетическим признакам. Аллотранспланация (аллотрансплантат) - пересадка тканей и органов между организмами одного биологического вида. Ксенотрансплантация (ксенотрансплантат) – пересадка тканей и органов между организмами разных биологических видов. Эксплантация (эксплантат) – пересадка небиологического материала. Комбинированная пересадка (комбинированный трансплантат). Остро стоят 2 проблемы: сохранение органов и тканей с их неизмененными свойствами. Другая проблема – преодоление трансплантационного иммунитета. Разные методы консервации. 1) Охлаждение (недолговременное). 2) Замораживание. 3) Лиофилизации. Заморозка может разорвать ткань, что приводит к гибели ткани. Но сперматозоиды способны жить. Состояние анабиоза некоторых животных. Кровь заменяют криопротекторами, после разморозки производят обратную замену. Метод лиофилизации – заморозка высушиванием в воздухе. Хранение замороженных людей. Существуют банки тканей, банки органов на научной основе. 2 проблема более сложна. Живые организмы многие миллионы лет были индивидуальными т.к. одни индивиды не смешивались с другими, поэтому преодолеть эту проблему весьма сложно, но паразиты не отторгаются организмом. В трансплантологии сначала считали, что отторжение происходит из-за различного макроскопического и микроскопического строения тканей. Однако теперь выяснилось, что реципиент и донор различаются набором специфических белков и антигенов. Аллогенные и ксеногенные органы и ткани, содержащие трансплантационные гены, в организме вызывают защитную реакцию – выработку антител. Защита направлена на уничтожение пересаженных органов и тканей у реципиента и состоит из нескольких сложнейших иммунно-биологических реакций. Человек ощущает эти процессы с 7 дня, максимум процесса достигается на 14-21 сутки. Преодоление тканевой несовместимости – работа хирургов, иммунологов, физиологов и других специалистов. Целое медицинское направление - иммунодепрессивная терапия – направлено на решение этой проблемы. Используют химические, физические и биологические факторы воздействия на организм реципиента. Физические методы – радиоактивное излучение, рентгеновские лучи. Химические методы – введение препаратов, снижающих иммунитет. Они сильно влияют на жизненно важные органы. Биологические методы – введение антитоксических сывороток, антибиотиков. Принцип действия - нейтрализация трансплантационных антител. Наиболее перспективный метод. В настоящее время пересаживают практически все: и органы, и ткани. История трансплантологии в России. 1933 – Ю.воронов – первая в мире пересадка почки. 1937 год – Демихов - первая в СССР пересадка сердца собаке. 1946 – Демихов – пересадил сердце и легкие собаке. 1948 – Демихов, Швековский – пересадка печени собаке. 1954 – Демихов пересадил вторую голову собаке. 1965 – Петровский – первая успешная пересадка почки. 1986 – Шумаков – первая в СССР пересадка сердца человеку.(1967 – Кристиан Бернард – ЮАР – успешная пересадка сердца человеку). 1990 – Ерамишанцев – первая в СССР пересадка печени человеку. В Воронеже существует центр по пересадке почек. В клинике Шарите в Германии ежегодно делаются 60-100 операций по пересадке печени. В 2005 году в Англии произведена успешная операция по пересадки печени от одного донора – ребенку и взрослому человеку. Несмотря на заслуги, трансплантология ограничена законодательством, кроме того, многие органы являются «дефицитными». 78.Биологические ритмы. Хронобиология и хрономедицина. 79.Биологическая эволюция. Современные теории эволюции. 80.Понятие о биологическом виде. Концепции вида. Реальность биологического вида. Структура и критерии вида. Вид как генетически изолированная система. 81.Популяция – элементарная единица вида. Основные характеристики популяции. Генетическая структура популяции. Закон Харди-Вайнберга: содержательное и математическое выражение. 82.Элементарные эволюционные факторы. Мутационный процесс, популяционные волны, изоляция, поток генов, генетико-автоматические процессы (дрейф генов), естественный отбор. Взаимодействие элементарных эволюционных факторов. 83.Естественный отбор. Формы естественного отбора. Творческая роль естественного отбора в эволюции. 84.Популяционная структура человечества. Дем. Изолят. Близкородственные и ассортативные браки. Особенности генофондов изолятов, их отличия от генофондов больших по размерам популяций людей. 85.Особенности действия элементарных эволюционных факторов в человеческих популяциях. Мутационный процесс и генетическая комбинаторика, волны численности, миграция населения, смешанные браки, гибридные популяции как поток генов, изоляция (территориальная и социальная), дрейф генов, естественный отбор. 86.Отбор в пользу и против гетерозигот. Примеры. 87.Генетический груз и его эволюционное значение. 88.Генетический полиморфизм: классификация. Адаптивный потенциал популяции человека 89.Генетический полиморфизм человечества: масштабы, факторы формирования. Значение генетического полиморфизма в прошлом, настоящем и будущем человечества (медико-биологический и социальный аспекты). 90.Макроэволюция. Её соотношение с микроэволюцией. Формы филогенеза (эволюции групп): филетическая и дивергентная эволюция, конвергентная эволюция и параллелизм. Примеры. 91.Макроэволюция. Типы (направления) эволюции групп. Арогенез и ароморфозы. Аллогенез и идиоадаптация. Примеры. 92.Макроэволюция. Биологический прогресс и биологический регресс, их основные критерии. Эмпирические правила эволюции групп. Примеры. 93.Индивидуальное и историческое развитие. Закон зародышевого сходства. Биогенетический закон. Рекапитуляция. Онтогенез – реализация генетической информации, происходящая на всех стадиях. Онтогенез – генетически контролируемый процесс. В ходе онтогенеза реализуется генотип и формируется фенотип. Онтогенез - индивидуальное развитие организма, совокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом от момента его зарождения до конца жизни. О. включает рост, т. е. увеличение массы тела, его размеров, дифференцировку. Термин "О." введён Э. Геккелем (1866) при формулировании им биогенетического закона. Первую попытку исторического обоснования О. сделал И. ф. Меккель. Проблема соотношения О. и филогенеза была поставлена Ч. Дарвином и разрабатывалась Ф. Мюллером,Э. Геккелем и др. Все связанные с изменением наследственности, новые в эволюционном отношении признаки возникают в О., но лишь те из них, которые способствуют лучшему приспособлению организма к условиям существования, сохраняются в процессе естественного отбора и передаются последующим поколениям, т. е. закрепляются в эволюции. Познание закономерностей, причин и факторов О. служит научной основой для отыскания средств влияния на развитие растений, животных и человека, что имеет важнейшее значение для практики растениеводства и животноводства, а также для медицины. Филогенез - историческое развитие организмов. Термин введён нем. эволюционистом Э. Геккелем в 1866. Основной задачей при изучении Ф. является реконструкция эволюционных преобразований животных, растений, микроорганизмов, установление на этой основе их происхождения и родственных связей между таксонами, к которым относятся изученные организмы. Для этой цели Э. Геккель разработал метод "тройного параллелизма", позволяющий путём сопоставления данных трёх наук – морфологии, эмбриологии и палеонтологии – восстановить ход исторического развития изучаемой систематической группы. Закон зародышевого сходства Исследователи начала XIX в. впервые стали обращать внимание на сходство стадий развития эмбрионов высших животных со ступенями усложнения организации, ведущими от низкоорганизованных форм к прогрессивным. Сопоставляя стадии развития зародышей разных видов и классов хордовых, К. Бэр сделал следующие выводы. 1. Эмбрионы животных одного типа на ранних стадиях развития сходны. 2. Они последовательно переходят в своем развитии от более общих признаков типа ко все более частным. В последнюю очередь развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, индивидуальные черты. 3. Эмбрионы разных представителей одного типа постепенно обособляются друг от друга. К. Бэр, не будучи эволюционистом, не мог связывать открытые им закономерности индивидуального развития с процессом филогенеза. Поэтому сделанные им обобщения имели значение не более чем эмпирических правил. Развитие эволюционной идеи в последующем позволило объяснить сходство ранних зародышей их историческим родством, а приобретение ими все более частных черт с постепенным обособлением друг от друга — действительным обособлением соответствующих классов, отрядов, семейств, родов и видов в процессе эволюции. Вскоре после открытия закона зародышевого сходства Ч. Дарвин показал, что этот закон свидетельствует об общности происхождения и единства начальных этапов эволюции в пределах типа. Биогенетический закон Геккеля-Мюллера: каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденного его предками или его видом (филогенез). Онтогенез — повторение филогенеза Сопоставляя онтогенез ракообразных с морфологией их вымерших предков, Ф. Мюллер сделал вывод о том, что ныне живущие ракообразные в своем развитии повторяют путь, пройденный их предками. Преобразование онтогенеза в эволюции, по мнению Ф. Мюллера, осуществляется благодаря его удлинению за счет добавления к нему дополнительных стадий или надставок. На основе этих наблюдений, а также изучения развития хордовых Э. Геккель (1866) сформулировал основной биогенетический закон, в соответствии с которым онтогенез представляет собой краткое и быстрое повторение филогенеза. Повторение структур, характерных для предков, в эмбриогенезе потомков названо рекапитуляциями. Рекапитулируют не только морфологические признаки — хорда, закладки жаберных щелей и жаберных дуг у всех хордовых, но и особенности биохимической организации и физиологии. Так, в эволюции позвоночных происходит постепенная утрата ферментов, необходимых для распада мочевой кислоты — продукта метаболизма пуринов. У большинства беспозвоночных конечный продукт распада мочевой кислоты — аммиак, у земноводных и рыб — мочевина, у многих пресмыкающихся — аллантоин, а у некоторых млекопитающих мочевая кислота вообще не расщепляется и выделяется с мочой. В эмбриогенезе млекопитающих и человека отмечены биохимические и физиологические рекапитуляции: выделение ранними зародышами аммиака, позже мочевины, затем аллантоина, а на последних стадиях развития — мочевой кислоты. Однако в онтогенезе высокоорганизованных организмов не всегда наблюдается строгое повторение стадий исторического развития, как это следует из биогенетического закона. Так, зародыш человека никогда не повторяет взрослых стадий рыб, земноводных, пресмыкающихся и млекопитающих, а сходен по ряду черт лишь с их зародышами. Ранние стадии развития сохраняют наибольшую консервативность, благодаря чему рекапитулируют более полно, чем поздние. Это связано с тем, что одним из наиболее важных механизмов интеграции ранних этапов эмбриогенеза является эмбриональная индукция, а структуры зародыша, формирующиеся в первую очередь, такие, как хорда, нервная трубка, глотка, кишка и сомиты, представляют собой организационные центры зародыша, от которых зависит весь ход развития. Генетическая основа рекапитуляции заключена в единстве механизмов генетического контроля развития, сохраняющемся на базе общих генов регуляции онтогенеза, которые достаются родственным группам организмов от общих предков. Рекапитуляция (от лат. recapitulatio – повторение) — понятие, используемое в биологии для обозначения повторения в индивидуальном развитии признаков, свойственных более ранней стадии эволюционного развития. 94.Онтогенез как основа филогенеза. Ценогенезы. Автономизация онтогенеза. Филэмбриогенезы. Учение А.Н.Северцова о филэмбриогенезах. Механизмы их возникновения. Гетерохронии и гетеротопии биологических структур в эволюции онтогенеза. Опираясь только на основной биогенетический закон, невозможно объяснить процесс эволюции: бесконечное повторение пройденного само по себе не рождает нового. Так как жизнь существует на Земле благодаря смене поколений конкретных организмов, эволюция ее протекает благодаря изменениям, происходящим в их онтогенезах. Эти изменения сводятся к тому, что конкретные онтогенезы отклоняются от пути, проложенного предковыми формами, и приобретают новые черты. К таким отклонениям относятся, например, ценогенезы — приспособления, возникающие у зародышей или личинок и адаптирующие их к особенностям среды обитания. У взрослых организмов ценогенезы не сохраняются. Примерами ценогенезов являются роговые образования во рту личинок бесхвостых земноводных, облегчающие им питание растительной пищей. В процессе метаморфоза у лягушонка они исчезают и пищеварительная система перестраивается для питания насекомыми и червями. К ценогенезам у амниот относят зародышевые оболочки, желточный мешок и аллантоис, а у плацентарных млекопитающих и человека — еще и плаценту с пуповиной. Ценогенезы, проявляясь только на ранних стадиях онтогенеза, не изменяют типа организации взрослого организма, но обеспечивают более высокую вероятность выживания потомства. Они могут сопровождаться при этом уменьшением плодовитости и удлинением зародышевого или личиночного периода, благодаря чему организм в постэмбриональном или постличиночном периоде развития оказывается более зрелым и активным. Возникнув и оказавшись полезными, ценогенезы будут воспроизводиться в последующих поколениях. Так, амнион, появившийся впервые у предков пресмыкающихся в каменноугольном периоде палеозойской эры, воспроизводится у всех позвоночных, развивающихся на суше, как у яйцекладущих — пресмыкающихся и птиц, так и у плацентарных млекопитающих. Другой тип филогенетически значимых преобразований филогенеза — филэмбриогенезы. Они представляют собой отклонения от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но имеющие адаптивное значение у взрослых форм. Так, закладки волосяного покрова появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но сам волосяной покров имеет значение только у взрослых организмов. Такие изменения онтогенеза, будучи полезными, закрепляются естественным отбором и воспроизводятся в последующих поколениях. В основе этих изменений лежат те же механизмы, которые обусловливают врожденные пороки развития: нарушение пролиферации клеток, их перемещения, адгезии, гибели или дифференцировки (см. § 8.2 и 9.3). Однако от пороков их так же, как и ценогенезы, отличает адаптивная ценность, т.е. полезность и закрепленность естественным отбором в филогенезе. В зависимости от того, на каких этапах эмбриогенеза и морфогенеза конкретных структур возникают изменения развития, имеющие значение филэмбриогенезов, различают три их типа. 1. Анаболии, или надставки, возникают после того, как орган практически завершил свое развитие, и выражаются в добавлении дополнительных стадий, изменяющих конечный результат. К анаболиям относят такие явления, как приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, неотличимый от других рыб, а также появление изгибов позвоночника, сращение швов в мозговом черепе, окончательное перераспределение кровеносных сосудов в организме млекопитающих и человека. 2. Девиации — уклонения, возникающие в процессе морфогенеза органа. Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих (см. § 14.4). В развитии легких у млекопитающих также обнаруживается рекапитуляция ранних стадий предков, позднее морфогенез идет по-новому (см. разд. 14.3.4). 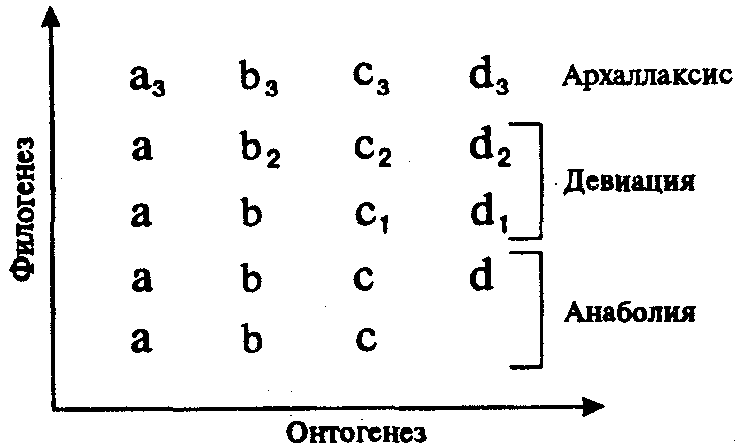 Рис. 13.9. Преобразования онто- и филогенеза в связи с возникающими филэмбриогенезами Буквами обозначены этапы онтогенеза, цифрами — филэмбриогенетические преобразования 3. Архаллаксисы — изменения, обнаруживающиеся на уровне зачатков и выражающиеся в нарушении их расчленения, ранних дифференцировок или в появлении принципиально новых закладок. Классическим примером архаллаксиса является развитие волос у млекопитающих, закладка которых наступает на очень ранних стадиях развития и с самого начала отличается от закладок других придатков кожи позвоночных (см. § 14.1). По типу архаллаксиса возникают хорда у примитивных бесчерепных, хрящевой позвоночник у хрящевых рыб (см. разд. 14.2.1.1), развиваются нефроны вторичной почки у пресмыкающихся (см. разд. 14.5.1). Ясно, что при эволюции за счет анаболии в онтогенезах потомков полностью реализуется основной биогенетический закон, т.е. происходят рекапитуляции всех предковых стадий развития. При девиациях ранние предковые стадии рекапитулируют, а более поздние заменяются развитием в новом направлении. Архаллаксисы полностью не допускают рекапитуляции в развитии данных структур, изменяя сами их зачатки. Если сопоставить схему филэмбриогенезов с таблицей К. Бэра (рис. 13.9), иллюстрирующей закон зародышевого сходства, то станет понятно, что Бэр уже был очень близок к открытию филэмбриогенезов, но отсутствие эволюционной идеи в его рассуждениях не позволило более чем на 100 лет опередить научную мысль. В эволюции онтогенеза наиболее часто встречаются анаболии как филэмбриогенезы, лишь в малой степени изменяющие целостный процесс развития. Девиации как нарушения морфогенетического процесса в эмбриогенезе часто отметаются естественным отбором и встречаются поэтому значительно реже. Наиболее редко в эволюции проявляются архаллаксисы в связи с тем, что они изменяют весь ход эмбриогенеза, и если такие изменения затрагивают зачатки жизненно важных органов или органов, имеющих значение эмбриональных организационных центров (см. разд. 8.2.6), то часто они оказываются несовместимыми с жизнью. В одной и той же филогенетической группе эволюция в разных системах органов может происходить за счет разных филэмбриогенезов. Так, в онтогенезе млекопитающих прослеживаются все этапы развития осевого скелета в подтипе позвоночных (анаболии), в развитии сердца рекапитулируют лишь ранние стадии (девиация), а в развитии придатков кожи рекапитуляции вообще отсутствуют (архаллаксис). Знание типов филэмбриогенезов в эволюции систем органов хордовых необходимо врачу для прогнозирования возможности возникновения у плодов и новорожденных врожденных пороков развития атавистической природы (см. разд. 13.3.4). Действительно, если в системе органов, эволюционирующей путем анаболии и девиаций, возможны атавистические пороки развития за счет рекапитуляции предковых состояний, то в случае архаллаксисов это исключается полностью. Кроме ценогенезов и филэмбриогенезов в эволюции онтогенеза могут обнаруживаться еще и отклонения времени закладки органов — гетерохронии — и места их развития — гетеротопии. Как первые, так и вторые приводят к изменению взаимосоответствия развивающихся структур и проходят жесткий контроль естественного отбора. Сохраняются лишь те гетерохронии и гетеротопии, которые оказываются полезными. Примерами таких адаптивных гетерохронии являются сдвиги во времени закладок наиболее жизненно важных органов в группах, эволюционирующих по типу арогенеза. Так, у млекопитающих, и в особенности у человека, дифференцировка переднего мозга существенно опережает развитие других его отделов. Гетеротопии приводят к формированию новых пространственных и функциональных связей между органами, обеспечивая в дальнейшем их совместную эволюцию. Так, сердце, располагающееся у рыб под глоткой, обеспечивает эффективное поступление крови в жаберные артерии для газообмена. Перемещаясь в загрудинную область у наземных позвоночных, оно развивается и функционирует уже в едином комплексе с новыми органами дыхания — легкими, выполняя и здесь в первую очередь функцию доставки крови к дыхательной системе для газообмена. Гетерохронии и гетеротопии в зависимости от того, на каких стадиях эмбриогенеза и морфогенеза органов они проявляются, могут быть расценены как филэмбриогенезы разных типов. Так, перемещение зачатков головного мозга, приводящее к его изгибу, характерному для амниот, и проявляющееся на начальных этапах его дифференцировки, является архаллаксисом, а гетеротопия семенника у человека из брюшной полости через паховый канал в мошонку, наблюдающаяся в конце эмбриогенеза после окончательного его формирования, — типичная анаболия. Иногда процессы гетеротопии, одинаковые по результатам, могут являться филэмбриогенезами разных типов. Например, у различных классов позвоночных очень часто встречается перемещение поясов конечностей. У многих групп рыб, ведущих придонный образ жизни, брюшные плавники (задние конечности) располагаются кпереди от грудных, а у млекопитающих и человека плечевой пояс и передние конечности в дефинитивном состоянии находятся значительно каудальнее места их первоначальной закладки. В связи с этим иннервация плечевого пояса у них осуществляется нервами, связанными не с грудными, а с шейными сегментами спинного мозга. У упомянутых выше рыб брюшные плавники иннервируются нервами не задних туловищных, а передних сегментов, расположенных кпереди от центров иннервации грудных плавников. Это свидетельствует о гетеротопии закладки плавников уже на стадии самых ранних зачатков, в то время как перемещение переднего пояса конечностей у человека происходит на более поздних этапах, когда иннервация их уже полностью осуществлена. Очевидно, в первом случае гетеротопия представляет собой архаллаксис, в то время как во втором — анаболию. Ценогенезы, филэмбриогенезы, а также гетеротопии и гетерохронии, оказавшись полезными, закрепляются в потомстве и воспроизводятся в последующих поколениях до тех пор, пока новые адаптивные изменения онтогенеза не вытеснят их, заменив собой. Благодаря этому онтогенез не только кратко повторяет эволюционный путь, пройденный предками, но и прокладывает новые направления филогенеза в будущем. |