ОБЩИЕ СВЕДЕНИЯ. Общие сведения
 Скачать 1.16 Mb. Скачать 1.16 Mb.
|

|
Нейрогипофиз Нейрогипофиз (см. рис. 37, 15; 38, 17) представляет собой производное дна воронки промежуточного мозга. Он находится в тесной морфологической и функциональной связи с гипоталамусом, в нем заканчиваются волокна гипоталамо-гипофизарного тракта, идущего от супраоптического и паравентрикулярного ядер гипоталамуса (см. рис. 37, 17, 20). Аденогипофиз (передняя доля) Аденогипофиз (передняя доля) развивается из эпителиального выпячивания (кармана Ратке) крыши кишечной трубки (см. рис. 37, 16; 38, 16). Передняя доля гипофиза имеет тесную сосудистую связь с гипоталамусом. Здесь артерии ветвятся на капилляры, образуя плотное сплетение в форме мантии на поверхности срединного возвышения. Капиллярные ветви этого сплетения образуют вены, достигающие передней доли гипофиза, здесь вены 75 вновь распадаются на капилляры, пронизывающие всю долю. Вся эта сложная система кровеносных сосудов носит название портальной. По ней в аденогипофиз из гипоталамуса поступают пептидные гормоны (либерины и статины), регулирующие синтез и секрецию гормонов аденогипофиза. Нейрогипофиз имеет собственную, не зависящую от портальной системы, систему кровоснабжения. В аденогипофизе секретируется два типа гормонов — эффекторные, т.е. реализующие свои свойства непосредственно в организме, и тропные — оказывающие регулирующее влияние на периферические железы внутренней секреции. Всего в аденогипофизе синтезируется шесть гормонов — гормон роста, пролактин, тиреотропин, адренокортикотропный гормон (АКТГ), фолликулостимулирующий гормон, лютенизирующий гормон. Фолликулостимулирующий и лютенизирующий гормоны объединяются в группу гонадотропных гормонов. За последние годы было установлено, что практически все биологически активные вещества, секретируемые нейронами гипоталамо-гипофизарной системы, имеют пептидную природу. Филогенез. Филогенез. В филогенезе структуры промежуточного мозга появляются и развиваются на разных его этапах. У низших позвоночных животных уже есть некоторые эпиталамические, метаталамические и гипоталамические структуры. У круглоротых таламус уже достаточно выражен и разделяется на дорсальную и вентральную части, включающие в себя несколько ядер. Гипоталамус является также выраженной структурой. У костистых рыб происходит усложнение в организации таламуса и дифференциация ряда ядерных групп. У амфибий и рептилий развиваются зрительные бугры. Они становятся специальными структурами и связывают средний мозг с конечным. У рептилий промежуточный мозг выполняет вместе с некоторыми структурами конечного мозга высшие интегративные функции. Дорсальный таламус у них отделяется от вентрального, на этом этапе выделяются четкие ядра с характерным строением и связями. В дорсальном таламусе их насчитывается девять, в вентральном — семь. В переднем отделе гипоталамуса у рептилий появляются зачатки супраоптического и паравентрикулярного ядер. У птиц дифференциация ядер гипоталамуса хорошо выражена, тогда как таламус усложняется незначительно. Особенно бурно идет развитие дорсального таламуса у млекопитающих, так как он выполняет функцию основного коллектора сенсорных путей к 76 коре больших полушарий. У млекопитающих наряду с развитием релейных (переключательных) ядер таламуса развиваются ассоциативные ядра. Происходит усиленное развитие таламуса и формируется таламонеокортикальная система интеграции. Онтогенез. Онтогенез. В онтогенезе структуры промежуточного мозга возникают из стенок второго мозгового пузыря (diencephalon) пятипузырной стадии развития головного мозга. Глазные пузырьки превращаются в глазные бокалы и глазные стебельки. Позже из них строятся сетчатка глаза и зрительные проводящие пути. Из боковых стенок второго пузыря формируются структуры дорсального (thalamus) и вентрального (subthalamus) таламусов. Верхней стенкой являются эпиталамические структуры, нижней — гипоталамус. Из верхней части задней стенки развиваются забугорные структуры (metathalamus). B этой части мозговой трубки все структуры образуются во время онтогенеза из крыловидной пластинки (пограничная борозда и основная пластинка заканчиваются на уровне среднемозгового пузыря). Поэтому здесь не возникают ни моторные (двигательные), ни вегетативные ядра, а следовательно, нет и соответствующих черепно-мозговых нервов. Все ядра промежуточного мозга являются либо сенсорными (переключательными), либо интегративными (ассоциативными). Имеются здесь и неспецифические ядра. Гипоталамус отделяется от соседних участков мозга на 2-м месяце внутриутробной жизни. Вслед за этим начинается формирование шести гипоталамических ядер, которые представляют собой скопление нейронов с определенными функциями. Дифференцировка входящих в них клеток продолжается до 6-го месяца внутриутробной жизни, а завершается еще позднее. В четырех из шести ядер, вырабатываются гормоны, которые по системе сосудов направляются в аденогипофиз. Гипоталамо-аденогипофизарную систему составляют супрахиазматическое ядро, вентромедиальное, дорсомедиальное и аркуатное ядра. Сосудистая система появляется на 14-й неделе в виде первых капиллярных петель, и ее формирование завершается к моменту рождения. Синтезируемые в этих ядрах регуляторные пептиды обнаруживаются в аденогипофизе уже на 10-й неделе развития плода. Однако, по некоторым наблюдениям, в первые три месяца внутриутробной жизни, а возможно, до первой половины беременности гипофиз не подчиняется контролю гипоталамуса. Это объясняется незрелостью нейросекреторных клеток и недостаточным развитием портальной системы сосудов. 77 Вазопрессин появляется в гипофизе плода на 15—17-й, а окситоцин — на 18—19-й неделях внутриутробного развития. К 6-му месяцу беременности содержание их значительно возрастает. Уже в этот период они принимают участие в регуляции жизнедеятельности плода. Установление гипоталамического контроля над эндокринными железами происходит к концу внутриутробного развития. Конечный мозг Конечный мозг (telencephalon) — самый большой отдел головного мозга. Он состоит из двух полушарий (hemispheria cerebri), которые разделены продольной мозговой щелью. Полушария сильно разрастаются и покрывают собой все остальные отделы головного мозга. Длина полушарий 170 мм, высота 120 мм.  Рис. 39. Схема взаимоотношений оболочек головного мозга: 1 — кость; 2 — твердая мозговая оболочка; 3 — паутинная оболочка; 4 — мягкая (сосудистая) оболочка; 5 — кровеносный сосуд мозга; 6 — подпаутинное пространство; 7 — субдуральное пространство; 8 — пахионовы грануляции 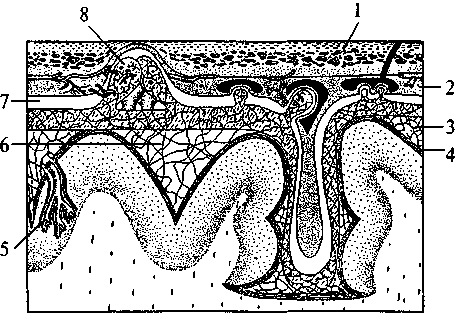 Полушария вместе со всем головным мозгом покрыты тремя мозговыми оболочками (рис. 39). Твердая мозговая оболочка складкой проникает глубоко между правым и левым полушариями и между затылочным полюсом полушарий и мозжечком. Паутинная оболочка входит во все борозды на поверхности. Под паутинной оболочкой находится подпаутинное пространство, заполненное спинномозговой жидкостью. Расширения подпаутинного пространства называются цистернами мозга. Мягкая мозговая оболочка по- а — схема поверхности мозга: 1 — цистерны мозга; б — схема сагиттального среза мозга: 1 — листки твердой оболочки мозга; 2 — паутинная оболочка; 3 — мягкая оболочка; 4 — задний рог бокового желудочка; 5 — нижний рог бокового желудочка; 6 — грануляции паутинной оболочки; 7 — отверстие Мажанди; 8 — центральный канал; 9 — отверстие Люшке; 10 — сосудистое сплетение IV желудочка; 11 — IV желудочек; 12 — Сильвиев водопровод; 13 — III желудочек; 14 — сосудистое сплетение III желудочка; 15 — передний рог бокового желудочка; 16 — Монроево отверстие; 17 — сосудистое сплетение бокового желудочка; 18 — 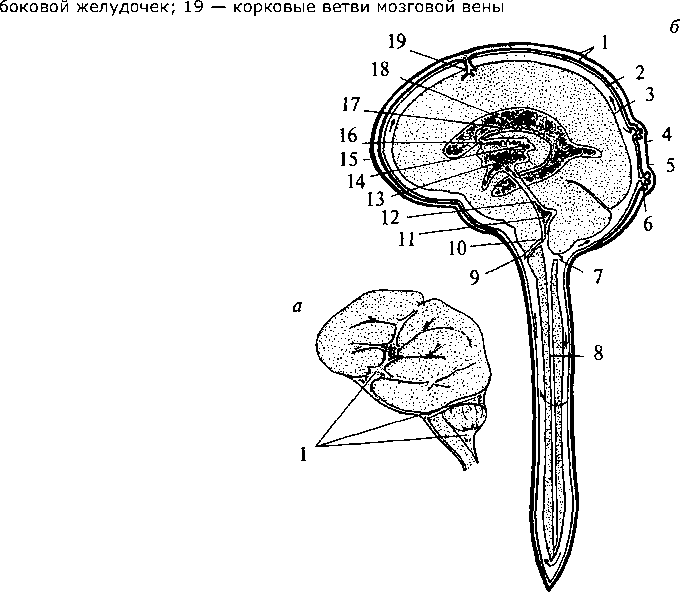 крывает всю поверхность полушарий (см. рис. 39). Подпаутинное пространство составляет функционально единое целое с венозной системой мозга и системой его желудочков (рис. 40). Каждое полушарие имеет свою щелевидную полость — боковой желудочек (рис. 40, 18). Условно желудочек левого полушария считается I, а правого — II. Вслед за ростом полушарий боковые желудочки разрастаются и принимают своеобразную форму. Кроме цент- 79 ральной части каждый желудочек имеет распространяющийся в лобную долю передний рог (рис. 40, 15), идущий в глубине затылочной доли задний рог (рис. 40, 4) и отходящий от него вниз и латерально нижний рог (рис. 40, 5), являющийся полостью височной доли. Боковые желудочки сообщаются с полостью III желудочка и имеют в своих стенках сосудистое сплетение. Медиальной стенкой каждого переднего рога является тонкая прозрачная пластинка. Правая и левая пластинки образуют между передними рогами общую прозрачную перегородку (septum pellucidum) (см. рис. 33, 2; 36, 3; 38, 2б). Расположение белого и серого вещества в полушариях сходно с таковым в мозжечке: в глубине белого вещества расположены ядра серого вещества, а вся поверхность полушарий покрыта серым веществом. Базальные ганглии В основании больших полушарий (нижняя стенка боковых желудочков) расположены ядра серого вещества — базальные ганглии. Они составляют примерно 3% от объема полушарий. Все базальные ганглии функционально объединены в две системы. Первая группа ядер представляет собой стриопаллидарную систему (рис. 41, 42, 43). К ним относятся: хвостатое ядро (nucleus caudatus), скорлупа (putamen) и бледный шар (globus pallidus). Скорлупа и хвостатое ядро имеют слоистую структуру, и поэтому их общее название — полосатое тело (corpus striatum). Бледный шар не имеет слоистости и выглядит светлее стриатума. Скорлупа и бледный шар объединены в чечевицеобразное ядро (nucleus lentiformis). Скорлупа образует наружный слой чечевицеобразного ядра, а бледный шар — внутренние его части. Бледный шар, в свою очередь, состоит из наружного и внутреннего члеников. Анатомически хвостатое ядро тесно связано с боковым желудочком. Расположенная впереди и медиально расширенная его часть — головка хвостатого ядра образует боковую стенку переднего рога желудочка, тело ядра — нижнюю стенку центральной части желудочка, а тонкий хвост — верхнюю стенку нижнего рога. Следуя форме бокового желудочка, хвостатое ядро охватывает дугой чечевицеобразное ядро (рис. 42, 1; 43, 1/). Друг от друга хвостатое и чечевицеобразное ядра отделены прослойкой белого вещества — частью внутренней капсулы (capsula interna). Другая часть внутренней капсулы отделяет чечевицеобразное ядро от нижележащего таламуса (рис. 43, 4). 80 Рис. 41. Полушария мозга на разных уровнях горизонтального разреза: (справа — ниже уровня дна бокового желудочка; слева — над дном бокового желудочка; IV желудочек мозга вскрыт сверху): 1 — головка хвостатого ядра; 2 — скорлупа; 3 — кора мозгового островка; 4 — бледный шар; 5 — ограда; 6 — хвост хвостатого ядра; 7 — ядра медиального коленчатого тела; 8 — нижний рог бокового желудочка; 9 — верхние ножки мозжечка; 10 — средние ножки мозжечка; 11 — нижние ножки мозжечка; 12 — мозговые полоски (волокна проводящих путей слуховой системы); 13 — треугольник подъязычного нерва; 14 — треугольник блуждающего нерва; 15 — задвижка; 16 — часть мозжечка; 17 — верхний мозговой парус; 18 — блоковый нерв; 19 — таламус; 20 — задняя ножка внутренней капсулы; 21 — конечная пластинка; 22 — колено внутренней капсулы; 23 — хвостатое ядро; 24 — кора мозгового островка; 25 — передняя ножка внутренней капсулы; 26 — передний рог бокового желудочка 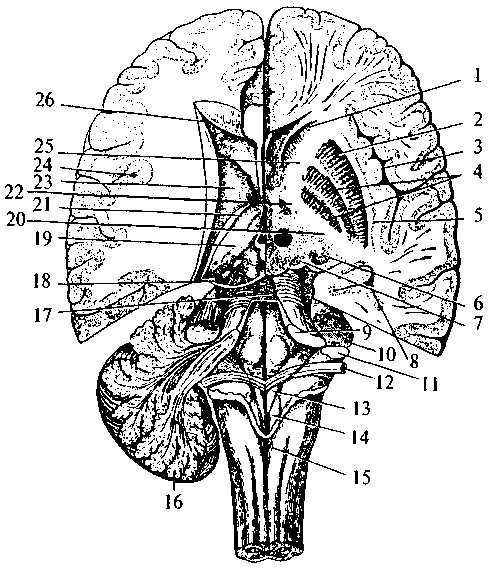 Таким образом, строение дна бокового желудочка (представляющего собой стриопаллидарную систему) схематично можно представить себе так: стенку самого желудочка образует слоистое хвостатое ядро, затем ниже идет прослойка белого вещества — Таким образом, строение дна бокового желудочка (представляющего собой стриопаллидарную систему) схематично можно представить себе так: стенку самого желудочка образует слоистое хвостатое ядро, затем ниже идет прослойка белого вещества —81 Рис. 42. Топография базальных ядер конечного мозга и стволовых структур (вид слева спереди): 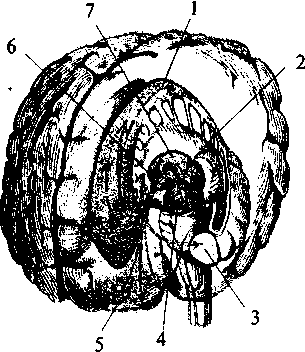 1 — хвостатое ядро; 2 — скорлупа; 3 — миндалина; 4 — черная субстанция; 5 — лобная кора; 6 — гипоталамус; 7 — таламус 1 — хвостатое ядро; 2 — скорлупа; 3 — миндалина; 4 — черная субстанция; 5 — лобная кора; 6 — гипоталамус; 7 — таламус |