ОБЩИЕ СВЕДЕНИЯ. Общие сведения
 Скачать 1.16 Mb. Скачать 1.16 Mb.
|

|
человека: 1 — область иннервации блуждающего нерва; 2 — область иннервации языкоглоточного нерва; 3 — область иннервации лицевого нерва; 4 — локализация грибовидных сосочков; 5 — локализация листовидных сосочков; 6 — локализация желобоватых сосочков 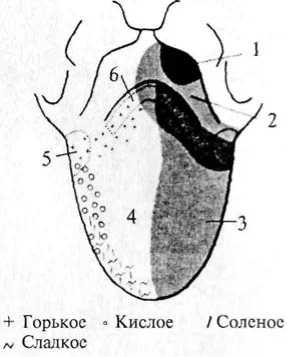 ную часть постцентральной извилины коры больших полушарий. Некоторые корковые нейроны этой зоны реагируют только на вещества с одним вкусовым качеством. Другие — и на механостимуляцию рецепторов ротовой полости, и на термостимуляцию. ную часть постцентральной извилины коры больших полушарий. Некоторые корковые нейроны этой зоны реагируют только на вещества с одним вкусовым качеством. Другие — и на механостимуляцию рецепторов ротовой полости, и на термостимуляцию.Филогенез. Филогенез. В филогенезе лишь у низших позвоночных происходит отделение вкусовой системы от общей хемочувствительности. У рыб уже имеется обособленная вкусовая система. Вкусовые почки располагаются не только во рту, но и на жабрах, на коже, на усиках, иногда на лучах плавников. У амфибий вкусовые рецепторы расположены только в ротовой полости и на языке. У рептилий и млекопитающих рецепторы расположены в основном на поверхности языка во вкусовых сосочках. Онтогенез. Онтогенез. Развитие вкусовой сенсорной системы начинается с 8-й недели эмбрионального развития. Сначала закладываются вкусовые почки, а к 4-му месяцу устанавливается их связь с черепно-мозговыми нервами. Завершается развитие вкусовой системы к 6-му месяцу эмбрионального развития. Обонятельная система Рецепторные клетки, воспринимающие запахи, расположены в обонятельном эпителии слизистой верхней носовой раковины. Обонятельная область слизистой имеет желтовато-коричневый цвет и занимает площадь около 10 см2. Обонятельные клетки слизистой (рис. 63, 4) соседствуют с опорными (рис. 63, 5) и базальными клетками. Опорные клетки вырабатывают слизь, в которую погружены волоски обонятельных булав (рис. 63, 3). Количество обонятельных клеток у человека около 30 млн. 115 Рис. 63. Строение обонятельного эпителия: 1 — комплексный волосок обонятельной клетки; 2 — волосок обонятельной клетки; 3 — обонятельная 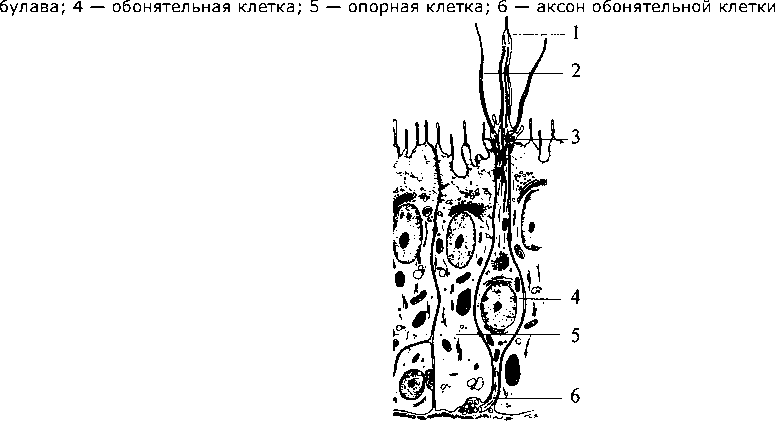 Обонятельные клетки являются первичными рецепторами. Их аксоны (рис. 63, 6) объединяются в 20—40 обонятельных нитей, которые через отверстия решетчатой кости проникают в полость черепа и направляются к обонятельной луковице (bulbus olfactorius) (см. рис. 55, 1). Обонятельные луковицы являются первичными обонятельными центрами. В них располагаются нейроны, на которых происходит переключение путей. От обонятельных луковиц начинаются обонятельные тракты (I пара черепно-мозговых нервов), по которым проходят аксоны нейронов обонятельных луковиц. Задние отделы обонятельных трактов уплощаются и расширяются, образуя обонятельные треугольники (см. рис. 55, 19). Часть аксонов (медиальная полоска обонятельного треугольника) (см. рис. 55, 20), дойдя до передней комиссуры больших полушарий, переходит по ней на противоположную сторону и в составе медиальной полоски тракта противоположной стороны возвращается к контрлатеральной луковице. Большая же часть волокон (латеральная полоска обонятельного треугольника) (см. рис. 55, 18) не перекрещиваясь идет к обонятельному бугорку, расположенному в области переднего продырявленного пространства, и к гиппокампу. Корковые проекции обонятельного анализатора расположены на вентральной стороне полушарий и относятся к древней коре. Обонятельный бугорок посылает свои аксоны к мамиллярным телам, миндалевидному комплексу и гиппокампу. Таким образом, обонятельные структуры имеют непосредственные связи с лимбической системой. Этим определяется особая роль обоняния в эмоциональных реакциях и процессах памяти. Филогенез. Орган обоняния позвоночных появляется в виде пары простых обонятельных ямок, выстланных изнутри чувствительным 116 эпителием, первоначально тесно связанным с передней частью нервной пластинки. У рыб необходимость смены воды в обонятельных ямках привела к появлению вводных и выводных отверстий — двух пар ноздрей. У наземных позвоночных выводная пара ноздрей перемещается внутрь ротовой полости и носит название хоан. Обонятельная полость становится частью дыхательных путей, однако обонятельный эпителий обособляется от эпителия дыхательных путей. Дальнейшее совершенствование обоняния у млекопитающих ведет к увеличению обонятельной поверхности путем появления выростов стенок обонятельной капсулы — носовых раковин. У млекопитающих с хорошо развитым обонянием их число достигает 5—8, а площадь обонятельного эпителия до 200 см2. Такие животные называются макросматиками. Человек, так же как и остальные приматы, пользуется обонянием в меньшей степени, и обонятельная сенсорная система у него развита слабее. Число внутренних носовых раковин у человека редуцируется до двух, хотя эмбрионально закладывается до пяти. Таким образом, он относится к микросматикам. И наконец, китообразные вообще не имеют обоняния и поэтому называются аносматиками. Конечный мозг у позвоночных первоначально выполняет только функцию анализа обонятельных стимулов. Но по мере развития животных он берет на себя множество других функций. У млекопитающих функцию анализа обонятельных стимулов выполняет небольшая поверхность коры, относящаяся к древней коре. У макросматиков — это грушевидная доля. У человека вследствие относительно большего развития неокортекса и уменьшения значения обоняния обонятельная кора занимает незначительную площадь поверхности. Онтогенез. Онтогенез. В онтогенезе у человека обонятельный эпителий закладывается уже на 4-й неделе развития в виде утолщений эктодермы в обонятельных ямках. С развитием носа обонятельный эпителий смещается в его полость. Аксоны рецепторных клеток прорастают к обонятельной луковице. Кожная рецепция Рецепторы, расположенные в коже, обеспечивают три типа чувствительности. Это тактильная, температурная и болевая чувствительность. 117  Рис. 64. Кожные рецепторы: 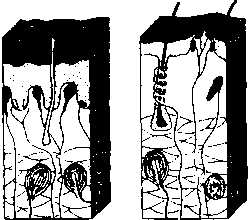 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини \ v t i 4 1 2 3 4 5 6 Тактильные ощущения (осязание) сигнализируют об особенностях нашего непосредственного окружения и обеспечиваются наличием в коже различных типов механорецепторов. Они имеют разное морфологическое строение и улавливают различные механические воздействия. Различают свободные и инкапсулированные рецепторы. Свободные — это нервные окончания, не имеющие вспомогательных структур и свободно расположенные в ткани. Инкапсулированные рецепторы включаются в комплекс со вспомогательными клетками. Среди инкапсулированных механорецепторов кожи различают: диски Меркеля, которые могут группироваться в тельца Пинкуса—Игго (рецепторы давления); окончания Руффини (также рецепторы давления); тельца Мейснера (рецепторы прикосновения); тельца Паччини (рецепторы смещения). Для участков кожи, покрытых волосками, характерны рецепторы волосяных фолликулов — свободные нервные окончания, обвивающие основание кожного волоска (рис. 64). Тонкость различения деталей тактильных стимулов зависит от плотности расположения рецепторов в коже. Так, на губах, языке, кончиках пальцев плотность расположения рецепторов наибольшая и соответственно различение тактильных особенностей наилучшее. Терморецепция обеспечивается специфическими тепловыми и Холодовыми рецепторами. Тепловые рецепторы являются свободными, холодовые — инкапсулированы. ПЛОТНОСТЬ Холодовых рецепторов в коже больше, чем тепловых. 118 Рис. 65. Соединение путей кожных и висцеральных рецепторов в спинном мозге: 1 — пучок Голля; 2 — пучок Бурдаха; 3 — задний корешок; 4 — передний корешок; 5 — спиноталамический тракт (проведение болевой чувствительности); 6 — двигательные аксоны; 7 — симпатические аксоны; 8 — передний рог; 9 — проприоспинальный путь; 10 — задний рог; И — висцерорецепторы; 12 — проприорецепторы; 13 — терморецепторы; 14 — ноцицепторы; 15 — механорецепторы 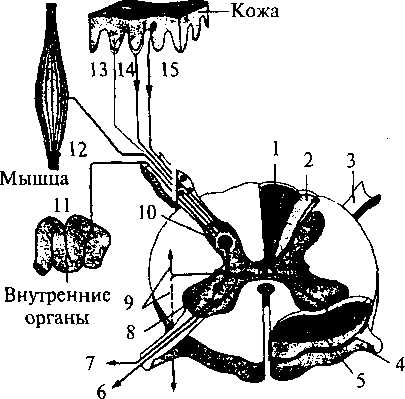 Болевая чувствительность (ноцицепция) Болевая чувствительность (ноцицепция) служит для распознавания потенциально опасных стимулов. Болевые рецепторы имеют самое плотное (по сравнению с тактильными и терморецепторами) расположение в коже (рис. 65). Однако распределены они не равномерно, образуя скопления — «болевые точки». Ноцицепторы являются свободными окончаниями. Они чувствительны к механическим, термическим и химическим стимулам, т.е. являются полимодальными. Все кожные рецепторы являются окончаниями псевдоуниполярных чувствительных нейронов, расположенных в спинномозговых ганглиях. По афферентным волокнам (дендритам) этих нейронов информация поступает сначала к телу нейрона, а затем по его аксону в задние рога соответствующего сегмента спинного мозга. В каждый задний корешок спинного мозга входят афференты, собирающие информацию с определенного участка кожи. Такой участок кожи называется дерматомом. Смежные дерматомы пере- 119 крываются, т.е. информация от одного участка кожи может поступать в смежные сегменты спинного мозга. Дерматомы лица и шеи иннервируются тройничным и лицевым нервами. Первичная обработка сигнала производится нейронами задних рогов сегмента спинного мозга (или соответствующими ядрами черепно-мозговых нервов) (рис. 65). От этих нейронов информация может поступать к мотонейронам и вегетативным (симпатическим) нейронам своего сегмента; далее короткими путями к соседним сегментам и, наконец, в протяженные восходящие пути спинного мозга (Голля и Бурдаха для тактильных и температурных воздействий (рис. 65, 1— 3) и спиноталамические для болевых воздействий). По трактам Голля и Бурдаха сигналы достигают одноименных ядер продолговатого мозга, затем переключаются в таламусе (вентробазальное ядро) и соматотопически проецируются в контрлатеральную постцентральную извилину. Спиноталамические пути, к которым присоединяются болевые афференты тройничного и лицевого нервов, переключаются в таламусе и проецируются также в постцентральную кору. Проприоцепция и интероцепция Кроме внешних (экстероцептивных) сенсорных систем, таких как зрение, слух и т.д, в организме выделяют «внутренние анализаторы», так называемые проприо- и интероцептивные системы. Эти системы обеспечивают поступление информации от двигательного аппарата и внутренних органов в высшие отделы нервной системы. Проприоцептивный анализатор Проприоцептивный анализатор воспринимает позу и движения нашего тела. Проприорецепторы расположены в суставах, сухожилиях и мышцах (рис. 65, 18). Выделяют несколько типов рецепторов, обеспечивающих проприорецепцию. В первую очередь это мышечные веретена поперечно-полосатых мышц, репетирующие степень растяжения мышечных волокон. Сухожильные органы обеспечивают информацию о степени натяжения сухожилий, а суставные механорецепторы — положении сустава и движении в нем. Рецептор является рабочим органом, периферической частью чувствительного нейрона. Тело нейрона расположено в спинномозговом ганглии, а периферический отросток (дендрит) псевдоуниполярного нейрона спинального ганглия заканчивается в тканях рецептором, аксон же входит в спинной мозг и участвует в формировании сенсорных путей. Группа мышц, иннервируемых 120 одним задним спинномозговым корешком, называется миотомом. Восходящие пути спинного мозга, несущие проприоцептивную информацию, — пути Голля и Бурдаха (см. рис. 65, 1-3). Эти пути переключаются в продолговатом мозге, затем информация поступает к мозжечку и через структуры среднего мозга приходит в таламус (вентробазальный комплекс), в котором происходит обработка практически всей проприоцептивной сенсорной информации, перед тем как она поступает в кору больших полушарий, в соответствующие рецептивные поля (первая и вторая соматосенсорные проекционные области коры). Следует отметить, что только часть проприоцептивной информации является осознаваемой, большая же ее часть не осознается. Интероцептивный анализатор Интероцептивный анализатор отражает состояние внутренней среды организма и его вегетативных органов. Информация от разнообразных интерорецепторов используется для бессознательных процессов регуляции, управляющих кровообращением, пищеварением, дыханием и т.д. Интерорецепторы висцеральной системы представлены хемо-, баро-, осмо-, термо- и другими типами рецепторов, передающих информацию через нервы вегетативной нервной системы (волокна блуждающего, чревного и тазового нервов) и восходящие пути спинного мозга. Блуждающий нерв передает информацию от рецепторов внутренних органов грудной и брюшной полости. Чревный нерв — от желудка, кишечника, брыжейки. Тазовый нерв — от органов малого таза Рецепторы соединительной ткани, сосудов и внутренних органов достаточно разнообразны. Встречаются рецепторы трех видов: свободные, несвободные и инкапсулированные. По определенным особенностям среди часто встречающихся рецепторов можно выделить клубочковые, древовидные и кустиковидные формы разной степени сложности. Клубочковыми по своей форме являются рецепторы давления. Кроме специфических по типу рецепции в периферической нервной системе и вегетативных ганглиях присутствуют разнообразные по форме рецепторы, выполняющие функции общей рецепции. Таким образом, спинной мозг является первичной структурой обработки сенсорной интероцептивной информации (рис. 65). Далее информация идет по спиноталамическому тракту к вентробазальному ядру таламуса. Из таламуса интероцептивная информация поступает в кору. |