ОБЩИЕ СВЕДЕНИЯ. Общие сведения
 Скачать 1.16 Mb. Скачать 1.16 Mb.
|

|
Рис. 15. Схема развития плаща конечного мозга (обозначен черным) в сравнении с остальными структурами мозга в ряду позвоночных: а — акула; б — ящерица; в — кролик; г — человек: 1 — обонятельная луковица; 2 — базальные ядра; 3 — промежуточный мозг; 4 — средний мозг; 5 — мозжечок; 6 — продолговатый мозг 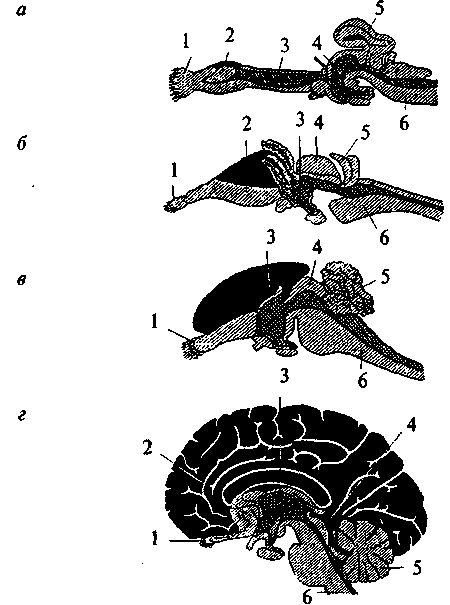 Для рептилий характерно значительное совершенствование передних отделов мозга. На поверхности плаща появляется серое вещество — кора. У высших представителей рептилий (крокодилы) в латеральных частях полушарий начинается закладка новой коры. Среди базальных ядер больших полушарий появляются но- 26 вые образования. В промежуточном мозге появляется зрительный бугор (таламус), имеющий специализированные ядра. Мозжечок хорошо развит и разбит на доли. Высшие интегративные функции выполняются промежуточным мозгом и базальными ядрами больших полушарий (рис. 15б, 2, 3). Совершенствование конечного мозга птиц идет по пути развития стриарных ядер. Корковые образования развиты слабо, новая кора отсутствует. Своими размерами выделяется мозжечок. Сенсорные и моторные функции распределены по отделам мозга так 27 же, как и у остальных позвоночных, но часть этих функций берет на себя стриатум конечного мозга. Высшие интегративные функции выполняются специфической для птиц структурой — добавочным гиперстриатумом. Развитие головного мозга млекопитающих пошло по пути увеличения относительной площади новой коры за счет развития складчатости плаща, наползания его на все остальные отделы головного мозга. Возникают связи новой коры с остальными отделами ЦНС и, соответственно, структуры, обеспечивающие их. В заднем мозгу появляется Варолиев мост, служащий для связи коры больших полушарий с мозжечком. Образуются средние ножки мозжечка, кроме того, в нем развиваются новые корковые структуры. В крыше среднего мозга появляется заднее двухолмие, с дорсальной стороны — ножки мозга. Продолговатый мозг приобретает пирамиды и оливы. Новая кора осуществляет почти все высшие сенсорные функции. За старой и древней корой остаются только обонятельные и висцеральные функции. У высших млекопитающих относительное представительство сенсорных функций уменьшается. Все большую поверхность коры занимают ассоциативные зоны коры (рис. 15г). Высшие интегративные функции у примитивных млекопитающих выполняют стриатум и кора, у высокоорганизованных — ассоциативные зоны новой коры. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ОНТОГЕНЕЗЕ Онтогенез, или индивидуальное развитие организма, делится на два периода: пренатальный (внутриутробный) и постнатальный (после рождения). Первый продолжается от момента зачатия и формирования зиготы до рождения; второй — от момента рождения и до смерти. Пренатальный период в свою очередь подразделяется на три периода: начальный, зародышевый и плодный. Начальный (предимплантационный) период у человека охватывает первую неделю развития (с момента оплодотворения до имплантации в слизистую оболочку матки). Зародышевый (предплодный, эмбриональный) период — от начала второй недели до конца восьмой недели (с момента имплантации до завершения закладки органов). Плодный (фетальный) период начинается с девятой недели и длится до рождения. В это время происходит усиленный рост организма. Постнатальный период онтогенеза подразделяют на одиннадцать периодов: 1-й — 10-й день — новорожденные; 10-й день — 1 год — грудной возраст; 1—3 года — раннее детство; 4—7 лет — первое детство; 8—12 лет — второе детство; 13—16 лет — подростковый период; 17—21 год — юношеский возраст; 22—35 лет — первый зрелый возраст; 36—60 лет — второй зрелый возраст; 61—74 года— пожилой возраст; с 75 лет — старческий возраст, после 90 лет — долгожители. Завершается онтогенез естественной смертью. Пренатальный период онтогенеза начинается с момента слияния мужских и женских половых клеток и образования зиготы. Зигота последовательно делится, образуя шаровидную бластулу. На стадии бластулы идет дальнейшее дробление и образование первичной полости — бластоцеля. Затем начинается процесс гаструляции, в результате которого происходит перемещение клеток различными способами в бласто- 29  А—А'— уровень поперечного среза; а — начальный этап погружения медуллярной пластинки и формирования нервной трубки: 1 — нервная трубка; 2 — ганглиозная пластина; 3 — сомит; б — завершение образования нервной трубки и погружение ее внутрь зародыша: 4 — эктодерма; 5 — центральный канал; 6 — белое вещество спинного мозга; 7 — серое вещество спинного мозга; 8 — закладка спинного мозга; 9 — закладка головного мозга 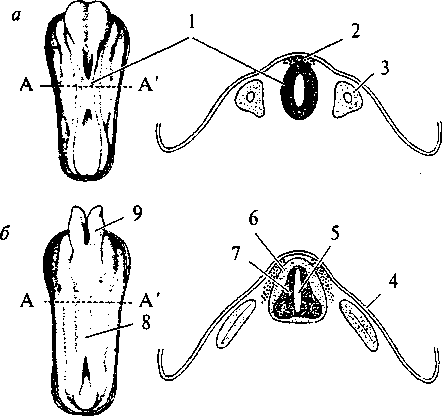 срезе): цель, с образованием двухслойного зародыша. Наружный слой клеток называется эктодерма, внутренний — энтодерма. Внутри образуется полость первичной кишки — гастроцель. Это стадия гаструлы. На стадии нейрулы образуются нервная трубка, хорда, сомиты и другие эмбриональные зачатки. Зачаток нервной системы начинает развиваться еще в конце стадии гаструлы. Клеточный материал эктодермы, расположенный на дорсальной поверхности зародыша, утолщается, образуя медуллярную пластинку (рис. 17, 2). Эта пластинка ограничивается с боков медуллярными валиками. Дробление клеток медуллярной пластинки (медуллобластов) и медуллярных валиков приводит к изгибанию пластинки в желоб, а затем к смыканию краев желоба и образованию медуллярной трубки (рис. 16а, 1). При соединении медуллярных валиков образуется ганглиозная пластина, которая затем делится на ганглиозные валики.  Рис. 17. Пренатальное развитие нервной системы человека: 1 — нервный гребень; 2 — нервная пластина; 3 — нервная трубка; 4 — эктодерма; 5 — средний мозг; 6 — спинной мозг; 7 — спинномозговые нервы; 8 — глазной пузырек; 9 — передний мозг; 10 — промежуточный мозг; 11 — мост; 12 — мозжечок; 13 — конечный мозг 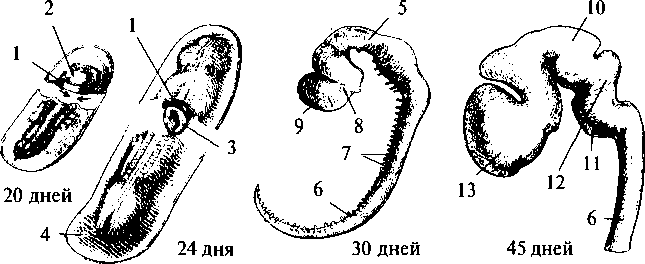 30 Одновременно происходит погружение нервной трубки внутрь зародыша (рис. 16в; 17, 3). Однородные первичные клетки стенки медуллярной трубки — медуллобласты — дифференцируются на первичные нервные клетки (нейробласты) и исходные клетки нейроглии (спонгиобласты). Клетки внутреннего, прилежащего к полости трубки, слоя медуллобластов превращаются в эпендимные, которые выстилают просвет полостей мозга. Все первичные клетки активно делятся, увеличивая толщину стенки мозговой трубки и уменьшая просвет нервного канала. Нейробласты дифференцируются на нейроны, спонгиобласты — на астроциты и олигодендроциты, эпендимные — на эпендимоциты (на этом этапе онтогенеза клетки эпендимы могут образовывать нейробласты и спонгиобласты). При дифференцировке нейробластов отростки удлиняются и превращаются в дендриты и аксон, которые на данном этапе лишены миелиновых оболочек. Миелинизация начинается с пятого месяца пренатального развития и полностью завершается лишь в возрасте 5—7 лет. На пятом же месяце появляются синапсы. Миелиновая оболочка формируется в пределах ЦНС олигодендроцитами, а в периферической нервной системе — Шванновскими клетками. В процессе эмбрионального развития формируются отростки и у клеток макроглии (астроцитов и олигодендроцитов). Клетки мик- 31 роглии образуются из мезенхимы и появляются в ЦНС вместе с прорастанием в нее кровеносных сосудов. Клетки ганглиозных валиков дифференцируются сначала в биполярные, а затем в псевдоуниполярные чувствительные нервные клетки, центральный отросток которых уходит в ЦНС, а периферический — к рецепторам других тканей и органов, образуя афферентную часть периферической соматической нервной системы. Эфферентная часть нервной системы состоит из аксонов мотонейронов вентральных отделов нервной трубки. В первые месяцы постнатального онтогенеза продолжается интенсивный рост аксонов и дендритов и резко возрастает количество синапсов в связи с развитием нейронных сетей. Эмбриогенез головного мозга начинается с развития в передней (ростральной) части мозговой трубки двух первичных мозговых пузырей, возникающих в результате неравномерного роста стенок нервной трубки (архэнцефалон и дейтерэнцефалон). Дейтерэнцефалон, как и задняя часть мозговой трубки (впоследствии спинной мозг), располагается над хордой. Архэнцефалон закладывается впереди нее. Затем в начале четвертой недели у зародыша дейтерэнцефалон делится на средний (mesencephalon) и ромбовидный (rhombencephalon) пузыри. А архэнцефалон превращается на этой (трехпузырной) стадии в передний мозговой пузырь (prosencephalon) (рис. 17, 9). В нижней части переднего мозга выпячиваются обонятельные лопасти (из них развиваются обонятельный эпителий носовой полости, обонятельные луковицы и тракты). Из дорсолатеральных стенок переднего мозгового пузыря выступают два глазных пузыря. В дальнейшем из них развиваются сетчатка глаз, зрительные нервы и тракты. На шестой неделе эмбрионального развития передний и ромбовидный пузыри делятся каждый на два и наступает пятипузырная стадия (рис. 17). Передний пузырь — конечный мозг — разделяется продольной щелью на два полушария. Полость также делится, образуя боковые желудочки. Мозговое вещество увеличивается неравномерно, и на поверхности полушарий образуются многочисленные складки — извилины, отделенные друг от друга более или менее глубокими бороздами и щелями (рис. 18). Каждое полушарие разделяется на четыре доли, в соответствие с этим полости боковых желудочков делятся также на 4 части: центральный отдел и три рога желудочка. Из мезенхимы, окружающей мозг зародыша, развиваются оболочки мозга. Серое вещество располагается и на периферии, образуя кору 32 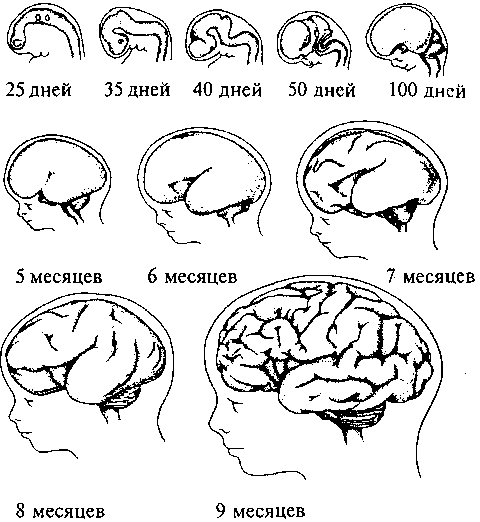 больших полушарий, и в основании полушарий, образуя подкорковые ядра. Задняя часть переднего пузыря остается неразделенной и называется теперь промежуточным мозгом (рис. 17, 10). Функционально и морфологически он связан с органом зрения. На стадии, когда границы с конечным мозгом слабо выражены, из базальной части боковых стенок образуются парные выросты — глазные пузыри (рис. 17, 5), которые соединяются с местом их происхождения при помощи глазных стебельков, впоследствии превращающихся в зрительные нервы. Наибольшей толщины достигают боковые стенки промежуточного мозга, которые преобразуются в зрительные бугры, или таламус. В соответствии с этим полость III желудочка превращается в узкую сагиттальную щель. В вентральной области (гипоталамус) образуется непарное выпячивание — воронка, из нижнего конца которой происходит задняя мозговая доля гипофиза — нейрогипофиз. 33 Третий мозговой пузырь превращается в средний мозг (рис. 17, 5), который развивается наиболее просто и отстает в росте. Стенки его утолщаются равномерно, а полость превращается в узкий канал — Сильвиев водопровод, соединяющий III и IV желудочки. Из дорсальной стенки развивается четверохолмие, а из вентральной — ножки среднего мозга. Ромбовидный мозг делится на задний и добавочный. Из заднего формируется мозжечок (рис. 17, 12) — сначала червь мозжечка, а затем полушария, а также мост (рис. 17, 11). Добавочный мозг превращается в продолговатый мозг. Стенки ромбовидного мозга утолщаются — как с боков, так и на дне, только крыша остается в виде тончайшей пластинки. Полость превращается в IV желудочек, который сообщается с Сильвиевым водопроводом и с центральным каналом спинного мозга. В результате неравномерного развития мозговых пузырей мозговая трубка начинает изгибаться (на уровне среднего мозга — теменной прогиб, в области заднего мозга — мостовой и в месте перехода добавочного мозга в спинной — затылочный прогиб). Теменной и затылочный прогибы обращены наружу, а мостовой — внутрь (рис. 17; 18). Структуры головного мозга, формирующиеся из первичного мозгового пузыря: средний, задний и добавочный мозг — составляют ствол головного мозга (truncus cerebri). Он является ростральным продолжением спинного мозга и имеет с ним общие черты строения. Проходящая по латеральным стенкам спинного мозга и стволового отдела головного мозга парная пограничная борозда (sulcus limitons) делит мозговую трубку на основную (вентральную) и крыловидную (дорзальную) пластинки. Из основной пластинки формируются моторные структуры (передние рога спинного мозга, двигательные ядра черепно-мозговых нервов). Над пограничной бороздой из крыловидной пластинки развиваются сенсорные структуры (задние рога спинного мозга, сенсорные ядра ствола мозга), в пределах самой пограничной борозды — центры вегетативной нервной системы. Производные архэнцефалона (telencephalon и diencephalon) создают подкорковые структуры и кору. Здесь нет основной пластинки (она заканчивается в среднем мозге), следовательно, и нет двигательных и вегетативных ядер. Весь передний мозг развивается из крыловидной пластинки, поэтому в нем имеются лишь сенсорные структуры (см. рис.18). 34 Постнатальный онтогенез нервной системы человека начинается с момента рождения ребенка. Головной мозг новорожденного весит 300—400 г. Вскоре после рождения прекращается образование из нейробластов новых нейронов, сами нейроны не делятся. Однако к восьмому месяцу после рождения вес мозга удваивается, а к 4—5 годам утраивается. Масса мозга растет в основном за счет увеличения количества отростков и их миелинизации. Максимального веса мозг мужчин достигает к 20—29 годам, а женщин к 15—19. После 50 лет мозг уплощается, вес его падает и в старости может уменьшиться на 100 г. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА Вегетативная нервная система регулирует внутренние процессы, обеспечивающие жизнь организма, такие как пищеварение, дыхание, сердечно-сосудистая деятельность. Центральные структуры вегетативной нервной системы расположены в головном и спинном мозге. В головном мозге это, прежде всего, гипоталамические центры, обеспечивающие постоянство внутренней среды организма, а также стволовые вегетативные ядра. В спинном мозге нейроны вегетативной нервной системы располагаются на границе между базальной и крыловидной пластинами, образуя боковые рога серого вещества. Периферические части вегетативной нервной системы состоят из ганглиев, представляющих собой скопления нервных клеток, лежащих вне пределов ЦНС, и волокон. Эфферентные волокна центральных структур вегетативной нервной системы выходят из ЦНС в составе смешанных черепно-мозговых или по передним корешкам спинномозговых нервов. Затем покидают общий нервный ствол и переключаются в ганглиях. Афферентные волокна заходят в ЦНС вместе с чувствительными соматическими волокнами через задние корешки спинного мозга или в составе черепно-мозговых нервов. Ганглии, в зависимости от местоположения, делятся на: паравертебральные (лежащие в непосредственной близости от позвоночника), превертебральные (удаленные от позвоночника), экстрамуральные (лежащие поблизости от иннервируемого органа) и интрамуральные (расположенные непосредственно в стенке органа). Волокна, подходящие к ганглию (преганглионарные), покрыты миелином, волокна, покидающие ганглий (постганглионарные), немиелинизированы и имеют серый цвет. В вегетативных ганглиях находятся афферентные, эфферентные и ассоциативные нейроны. Для этих ганглиев характерно, что 36 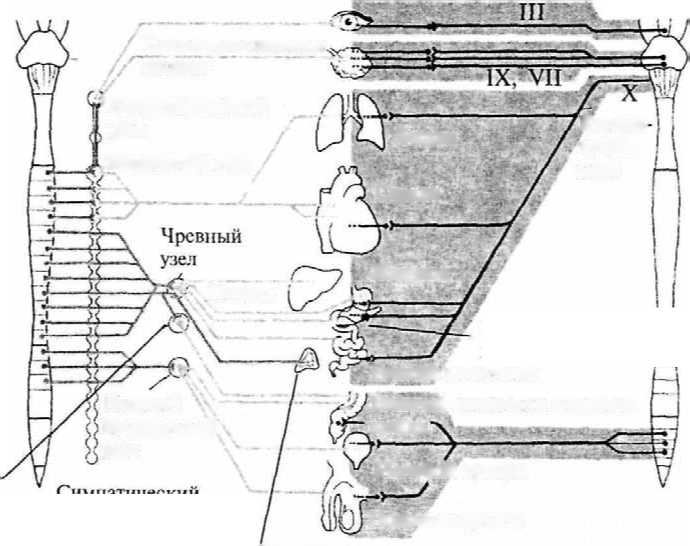     |