ОБЩИЕ СВЕДЕНИЯ. Общие сведения
 Скачать 1.16 Mb. Скачать 1.16 Mb.
|

|
Восходящие пути Восходящие пути состоят из аксонов клеток спинальных ганглиев и пучковых клеток серого вещества. К восходящим путям относятся: тонкий (нежный) пучок Голля (рис. 24, 18); клиновидный пучок Бурдаха (рис. 24, 17); Эти пути сформированы из отростков клеток спинальных ганглиев (тонкого — от 19 нижних сегментов, а клиновидного от 12 верхних сегментов спинного мозга). Их волокна вступают в спинной мозг через задние корешки и отдают коллатерали нейронам серого вещества. Сами же аксоны достигают одноименных ядер продолговатого мозга. вентральный и латеральный спиноталамические пути (рис. 24, 13); Они начинаются от чувствительных клеток спинномозговых ганглиев, которые переключаются на пучковых клетках задних рогов серого вещества. Аксоны этих клеток переходят по серому веще- 48 ству на противоположную сторону и достигают переключательных ядер таламуса. дорсальный спиномозжечковый путь Флексинга (рис. 24, 76); Этот путь начинается нейронами спинно-мозговых узлов, переключается на клетках ядра Кларка. Аксоны не переходят на противоположную сторону и, достигая продолговатого мозга, через нижнюю ножку мозжечка вступают в ипсилатеральную (той же стороны) половину мозжечка. вентральный спиномозжечковый путь Говерса (рис. 24,11); Он также начинается нейронами спинномозговых узлов, затем переключается на клетки ядра промежуточной зоны. Аксоны переходят на противоположную сторону и поднимаются до верхних отделов ствола, где вновь перекрещиваются и вступают в ипсилатеральную половину мозжечка по его верхним ножкам. К нисходящим путям относятся: К нисходящим путям относятся: латеральный и вентральный кортикоспинальные (пирамидные) пути (рис. 24, 75); Эти пути начинаются от пирамидных нейронов нижних слоев моторной зоны коры. Они проходят через белое вещество больших полушарий, основание ножек среднего мозга, по вентральным отделам Варолиева моста и продолговатого мозга в спинной мозг. Латеральный путь перекрещивается в нижней части пирамид продолговатого мозга и заканчивается на нейронах основания заднего рога. Вентральный путь пересекает пирамиды продолговатого мозга, не перекрещиваясь. Перед вступлением в передний рог серого вещества соответствующего сегмента спинного мозга волокна этого пути переходят на противоположную сторону и заканчиваются на мотонейронах передних рогов контрлатеральной стороны. руброспинальный путь (рис. 24, 14); Начинается от красных ядер среднего мозга, перекрещивается на уровне среднего мозга. Волокна заканчиваются на нейронах промежуточной зоны серого вещества спинного мозга. тектоспинальный путь (рис. 24, 7); Берет начало от клеток четверохолмия среднего мозга и достигает мотонейронов передних рогов. оливоспинальный путь (рис. 24, 10); Этот путь образован аксонами клеток олив продолговатого мозга, которые достигают мотонейронов спинного мозга. 49 вестибулоспинальные пути (рис. 24, 9, 12); Начинаются от вестибулярных ядер продолговатого мозга и заканчиваются на клетках передних рогов. ретикулоспинальный путь. Связывает ретикулярную формацию ствола головного мозга со спинным мозгом. Помимо этих путей имеются внутренние, локальные пути, соединяющие сегменты различных уровней спинного мозга между собой. Филогенез. В филогенезе спинного мозга отмечается вариабельность количества сегментов у представителей различных классов и видов. Но, являясь филогенетически древней структурой, спинной мозг мало изменяется в ходе эволюции. Пирамидные пути появляются только у млекопитающих в связи с развитием коры больших полушарий. Удельный вес спинного мозга по отношению к общей массе ЦНС в ходе филогенеза уменьшается за счет увеличения массы головного мозга. Онтогенез. В онтогенезе спинной мозг формируется из заднего отдела медуллярной трубки. На первых стадиях развития нервная трубка на всем протяжении состоит из трех слоев: наружного (краевого), среднего (мантийного), внутреннего (эпендимного). Эпендимные клетки выстилают внутренние стенки полости мозговой трубки. Клетки мантийного слоя преобразуются в нейробласты и спонгиобласты. Стенки трубки утолщаются неравномерно. Основной прирост нервной ткани идет в ее боковых частях. Нервные и глиальные клетки боковых стенок спинномозговой трубки образуют две пластинки: крыловидную и базальную. Крыловидная пластинка располагается дорсолатерально вдоль дорсальной части нервной трубки, а основная, или базальная, лежит вентролатерально. Правая и левая половины пластинок соединены с дорсальной стороны тонкой крышей, а с вентральной стороны — дном. Причем и дно, и крыша расположены в глубине, около центрального канала. Отделяются пластинки друг от друга боковыми пограничными бороздами. Клетки крыловидной пластинки формируют чувствительные, сенсорные структуры задних отделов спинного мозга (задние рога серого вещества). Базальная пластинка дает начало клеткам передних рогов — моторным структурам. Клетки, расположенные вблизи пограничных борозд, образуют вегетативные центры спинного мозга. Полость нервной трубки сужается из-за роста стенок спинного мозга и превращается в центральный спинномозговой канал. Возникают борозды, которые делят спинной мозг на 50 правую и левую половины. Отростки нейронов спинного и головного мозга прорастают, покрываются миелином и формируют белое вещество спинного мозга. Головной мозг Продолговатый мозг В полости черепа спинной мозг без резких морфологических изменений переходит в продолговатый мозг (medulla oblongata). Полость центрального канала на уровне продолговатого мозга расширена и образует нижнюю часть IV желудочка мозга (ventriculus qvartus), имеющего ромбовидную форму. Крыловидные пластинки дорсальной части мозговой трубки расходятся в стороны. Крыша мозга истончена, в ней располагается сосудистое сплетение IV желудочка. Поэтому продолговатый мозг имеет конусовидную форму. Узкий его конец является продолжением спинного мозга, а широкий направлен в сторону моста. Нижней границей продолговатого мозга являются верхние корешки шейного отдела спинного мозга, а верхняя граница проходит с дорсальной стороны по мозговым полоскам (волокна проводящих путей слуховой системы), идущим поперек дна IV желудочка в самой широкой его части, а с вентральной стороны — по нижнему краю моста. На дорсальной поверхности продолговатого мозга хорошо заметна полость треугольной формы - нижняя часть ромбовидной ямки дна IV желудочка. Срединная борозда ямки каудально (вершина треугольника) переходит в центральный спинномозговой канал. Часть крыши мозга, прикрывающая это место (точка расхождения крыловидных пластинок), называется задвижка (obex) (рис. 25, 18). Крышу мозга над продолговатым мозгом образуют сосудистое сплетение IV желудочка и нижний мозговой парус (см. рис. 33, 19). Через три отверстия в крыше мозга — срединную апертуру (apertum medialis ventriculi quarti), или отверстие Мажанди (см. рис. 40, 7), и два латеральные (aperturae laterales ventriculi quarti), или отверстия Люшке (см. рис. 40, 9), полость IV желудочка сообщается с подпаутинным пространством. Отверстие Мажанди больше латеральных, находится в задней части нижнего мозгового паруса и открывается в расширенную часть подпаутинного пространства — цистерну мозжечка и спинного мозга. Отверстия Люшке расположены латерально в самой широкой части ромбовидной ямки. Таким обра- 51 Рис. 25. Ствол мозга с дорсальной стороны. Мозжечок убран: 1 — язычок мозжечка; 2 — верхний мозговой парус; 3 — верхняя ямка; 4 — верхняя ножка мозжечка; 5 — средняя ножка мозжечка; 6 — нижняя ножка мозжечка; 7 — слуховой бугорок; 8 — мозговые полоски; 9 — треугольник подъязычного нерва; 10 — бугорок нежного ядра; 11 — боковой канатик; 12 — клиновидный канатик; 13 — нежный канатик; 14 — задняя срединная борозда; 15 — задняя промежуточная борозда; 16 — задняя боковая борозда; 17 — бугорок клиновидного ядра; 18 — задвижка; 19 — треугольник блуждающего 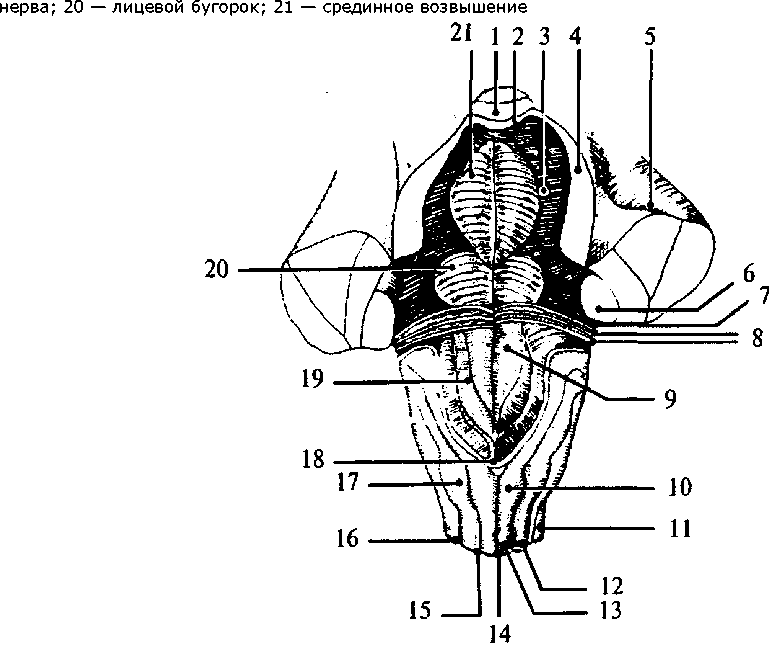 зом, на уровне продолговатого мозга ликвор желудочков мозга и спинномозговая жидкость объединяются в единую систему. С дорсальной поверхности продолговатого мозга от спинного мозга вдоль срединно-задней борозды поднимаются канатики нежного пучка (рис. 25, 13), латеральнее идут клиновидные канатики (рис. 25, 12). Канатики заканчиваются бугорками ядер Голля (нежного) и Бурдаха (клиновидного) (рис. 25, 10, 17) на уровне нижнего угла ромбовидной ямки. От бугорков Голля и Бурдаха рострально идут веревчатые тела (corpus restiforme), которые являются латеральными стенками IV желудочка. Они переходят в нижние ножки мозжечка (pedunculus cerebellaris inferior) (рис. 25, 6). 52  III — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI — отводящий нерв; VII — лицевой нерв; VIII — предверно-улитковый нерв (слуховой); IX — языкоглоточный нерв; X — блуждающий нерв; XI — добавочный нерв; XII — подъязычный нерв; 1 — ножка мозга (средний мозг); 2 — перекрест пирамид (верхний шейный отдел спинного мозга); 3 — боковой канатик; 4 — передняя боковая борозда; 5 — олива; 6 — пирамида продолговатого мозга; 7 — мост 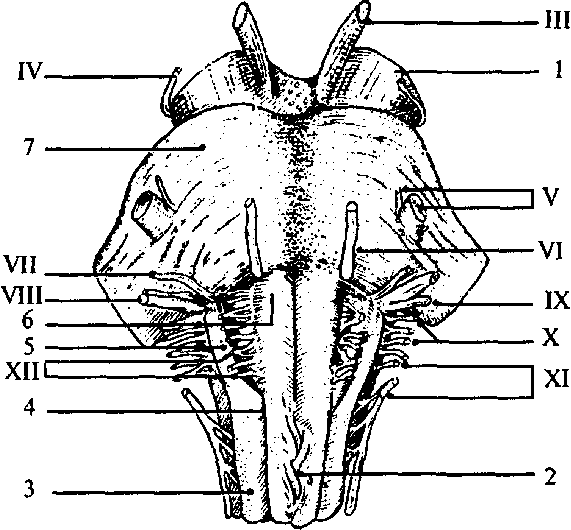 выхода черепно-мозговых нервов): На вентральной поверхности продолговатого мозга нисходящие кортикоспинальные проводящие пути образуют расширяющиеся валики — пирамиды (pyramis) (рис. 26, 6), лежащие по обе стороны от передней щели. Латеральнее от них расположены вздутия, называемые оливами (oliva), которым внутри соответствуют ядра серого вещества (рис. 26, 5). От продолговатого мозга отходят IX-XII пары черепно-мозговых нервов (рис. 26; 27): — языкоглоточный нерв (я. glossopharingeus), — блуждающий нерв (п. vagus), — добавочный нерв (п. accessorius), — подъязычный нерв (п. hypoglossus). 53 выхода черепно-мозговых нервов): — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI отводящий нерв; VII — лицевой нерв; VIII — предверно-улитковый нерв (слуховой); IX — языкоглоточный нерв; X — блуждающий нерв; XI — добавочный нерв; XII — подъязычный нерв; 1 — верхний бугорок четверохолмия (средний мозг); 2 — нижний бугорок четверохолмия (средний мозг); 3 — верхняя ножка мозжечка; 4 — средняя ножка мозжечка; 5 — слуховой бугорок; 6 — нижняя ножка мозжечка; 7 — задняя боковая борозда; 8 — бугорок нежного ядра; 9 — бугорок клиновидного ядра; 10 — пирамида; 11 — передняя боковая борозда; 12 — олива; 13 — мост; 14 — ножки мозга (средний мозг); 15 — ручка нижнего бугорка (средний мозг)  Языкоглоточный нерв Языкоглоточный нервЯзыкоглоточный нерв включает чувствительные, двигательные и парасимпатические ветви. Он выходит из продолговатого мозга через латеральную щель. Блуждающий нерв Блуждающий нерв является самым значительным нервом парасимпатического отдела нервной системы. Это смешанный нерв. Добавочный и подъязычный нервы Добавочный и подъязычный нервы чисто двигательные нервы. Добавочный нерв имеет веточки, выходящие из шейных сегментов спинного мозга. Структурная организация продолговатого мозга отличается от организации спинного мозга. Скопления клеток — ядра продолговатого мозга — перемежаются с пучками волокон, которые не всегда имеют поверхностное расположение. Чувствительные (сенсорные) ядра черепно-мозговых нервов располагаются дорсолатерально, двигательные (моторные) — вентрально, вегетативные ядра за- 54 Левая половина — схема расположения ядер (обозначены светлым) и проводящих путей (обозначены темным): 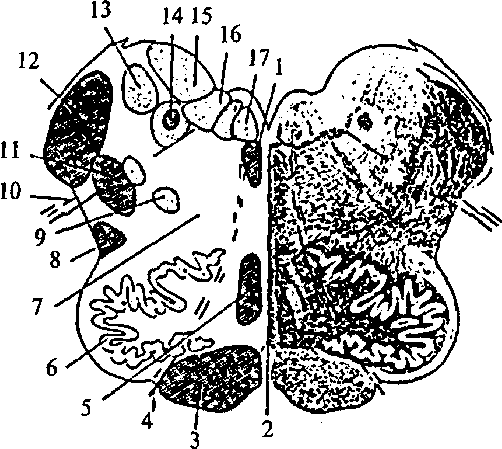 1 — задний продольный пучок; 2 — шов; 3 — пирамида; 4 — корешок подъязычного нерва; 5 — медиальная петля (осязательный путь); 6 — ядро оливы; 7 — ретикулярная формация; 8 — спиноталамический тракт; 9 — двойное ядро; 10— корешок блуждающего нерва; 11 — дорсальный продольный пучок; 12 — нижние ножки мозжечка; 13 — ядро Дейтерса; 14 — одиночный (солитарный) тракт и его ядро; 15 — верхнее вестибулярное ядро; 16 — дорсальное ядро блуждающего нерва; 17 — ядро подъязычного нерва 1 — задний продольный пучок; 2 — шов; 3 — пирамида; 4 — корешок подъязычного нерва; 5 — медиальная петля (осязательный путь); 6 — ядро оливы; 7 — ретикулярная формация; 8 — спиноталамический тракт; 9 — двойное ядро; 10— корешок блуждающего нерва; 11 — дорсальный продольный пучок; 12 — нижние ножки мозжечка; 13 — ядро Дейтерса; 14 — одиночный (солитарный) тракт и его ядро; 15 — верхнее вестибулярное ядро; 16 — дорсальное ядро блуждающего нерва; 17 — ядро подъязычного нерванимают промежуточное положение. Кроме ядер черепно-мозговых нервов в продолговатом мозге расположены ядра, на которых переключается сенсорная информация (ядра олив, ядра Голля и Бурдаха), и ядра ретикулярной формации, а также дыхательный, сосудодвигательный и пищеварительные центры (рис. 28). В продолговатом мозге и других стволовых отделах мозга (Варолиевом мосту и среднем мозге) расположена особая клеточная структура — ретикулярная формация (рис. 28, 7). В функциональном отношении ретикулярная формация ствола представляет собой единую структуру. Название этой структуры отражает ранние представления о том, что отдельные нейроны ретикулярной формации имеют обширные связи друг с другом и образуют нечто подобное нейропилю, в котором возбуждение распространяется диффузно, 55 подобно тому, как это происходит в нервной системе кишечнополостных. Однако в дальнейшем эти представления не подтвердились. Четких границ между отдельными ретикулярными и неретикулярными группами клеток не установлено, однако выделено до 98 ядерных групп, относящихся к ретикулярной формации. Основными ядрами являются: ядра шва и гигантоклеточное ядро продолговатого мозга, центральное и ретикулярное ядра моста. Клетки ретикулярной формации очень разнообразны по форме и величине. Для них характерно наличие значительно разветвленного дендритного дерева и длинных аксонов. Афферентные входы ретикулярная формация получает как по коллатералям восходящих (сенсорных) путей, так и от вышележащих структур, в том числе от коры больших полушарий и мозжечка. Таким образом, ретикулярная формация интегрирует влияние большого числа мозговых структур. В свою очередь сама она оказывает влияние как на вышележащие, так и на нижележащие структуры. Нисходящие и восходящие волокна ретикулярной формации покидают ядра на всем ее протяжении, не имея четких пространственных разграничений. Кроме того, существуют аксоны, которые Т-образно ветвятся. Одна из ветвей идет вниз, а другая — в верхние отделы головного мозга. Нисходящие волокна образуют ретикулоспинальный тракт. Через него ретикулярная формация оказывает влияние как на двигательную деятельность спинного мозга (осуществление спинальных рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная, пищеварительная функции). Ретикулярная формация воздействует на соматические и вегетативные центры в двух противоположных направлениях: торможения и возбуждения. Восходящие влияния ретикулярной формации направлены на регуляцию деятельности коры больших полушарий. Большинство ретикулярных волокон достигает коры через переключение в неспецифических ядрах таламуса. Действие восходящих ретикулярных влияний заключается в широкой активации корковых структур. Торможение активности ретикулярной формации ведет к наступлению сна, активация ее приводит к реакции пробуждения. Характерной особенностью нейронов ретикулярной формации является их высокая чувствительность к химическим факторам. Такие факторы, как уровень углекислого газа и кислорода, содержание адреналина, ацетилхолина и серотонина, относительно небольшие концентрации фармакологических веществ, изменяют актив- 56 ность нейронов ретикулярной формации, а вместе с этим и ее влияние на кору больших полушарий, соматические и вегетативные рефлексы. |