М1.+Урок+6_Доп.+материалы. Онлайнкурс Учимся легко нейропсихология в детском саду и школе Модуль Урок Дополнительные материалы
 Скачать 307.34 Kb. Скачать 307.34 Kb.
|

|
Развитие структуры коры передней центральной извилины в филогенезе
Решающее значение в подготовке двигательных импульсов имеют вторичные и третичные зоны, подчиняющиеся тем же принципам иерархического строения и убывающей специфичности, которые мы сформулировали, обсуждая принципы функциональной организации второго блока мозга. Основным отличием здесь является тот факт, что в третьем блоке процессы идут в нисходящем направлении, начинаясь в третичных зонах, где формируются двигательные планы и программы, переходя затем к аппаратам первичной двигательной зоны, которая посылает подготовленные двигательные импульсы на периферию. Другое отличие третьего блока коры от второго заключается в том, что этот блок не содержит модально-специфических зон. Премоторные отделы третьего блока мозга можно отнести к вторичным отделам коры. Префронтальные отделы мозга состоят из клеток третичных зон коры. Они играют решающую роль в формировании намерений и программ, в регуляции и контроле наиболее сложных форм поведения человека. Префронтальная область мозга имеет богатейшую систему связей с нижележащими отделами мозга и соответствующими отделами ретикулярной формации, практически со всеми остальными отделами коры. Благодаря двустороннему характеру этих связей префронтальные отделы коры находятся в особенно выгодном положении, как для вторичной переработки информации, так и для регуляции всех структур мозга. Влияние префронтальной коры на высшие формы процессов активации, регулируемые с помощью речи, было подробно изучено Е.Д.Хомской и ее сотрудниками. Следует отметить тот факт, что префронтальные отделы мозговой коры созревают на поздних этапах онтогенеза. Новейшие данные, относящиеся к динамике развития префронтальных отделов лобной коры, иллюстрирует рис. 4. Темп роста площади лобных областей мозга резко повышается к 3,5-4 годам; второй скачок приходится на возраст 7 - 8 лет. 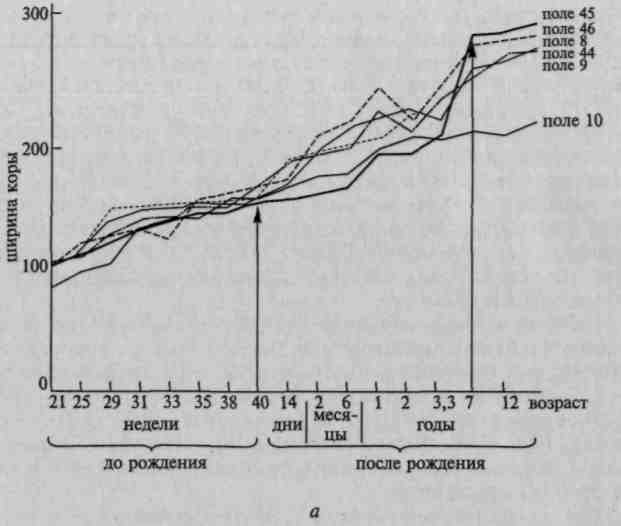 Рис. 4. Темп роста площади лобных областей мозга в онтогенезе: а — общая схема роста полей лобной области до и после рождения; б — темп роста поверхности и размеров клеток лобной коры (по данным Московского института мозга) В многочисленных исследованиях отмечаются обширные связи лобных долей как с затылочными, височными, теменными областями, так и с лимбическими отделами коры (Прибрам, 1961, 1971; Розе, Вулси, 1949; Загер, 1962, 1965, 1968; Наута, 1964; Пандуа, Купере, 1968, 1969). В отличие от третичных зон задних отделов мозга третичные отделы лобных долей фактически надстроены над всеми отделами мозговой коры, выполняя, таким образом, гораздо более универсальную функцию общей регуляции поведения, чем та, которую осуществляет второй блок мозга. Образования третьего блока вносят огромный вклад в общую организацию поведения человека. Уже ранние наблюдения над животными, лишенными лобных долей мозга, показали, насколько глубоко изменяется поведение животного. Как указывал И.П.Павлов, у такого животного нельзя отметить каких-либо нарушений в работе отдельных органов чувств: зрительный, слуховой и кинестетический анализы остаются у него сохранными, однако осмысленное, целенаправленное поведение глубоко страдает. В. М. Бехтерев высказал предположение о том, что лобные доли мозга играют важную роль в «правильной оценке внешних впечатлений и целесообразном, направленном выборе движений, сообразно с упомянутой оценкой», обеспечивая, таким образом, «психорегуляторную деятельность». В наши дни П. К. Анохин высказал предположение, что лобные доли мозга играют существенную роль в «синтезе обстановочных сигналов», обеспечивая «предварительную, предпусковую афферентацию» поведения. Разрушение префронтальной коры приводит к глубокому нарушению сложных программ поведения и к выраженному растормаживанию непосредственных реакций на побочные раздражители, в результате чего выполнение сложных программ поведения становится невозможным. Многочисленные наблюдения показывают, что лобные доли осуществляют не только функцию синтеза внешних раздражителей, подготовки к действию и формирования программ, но и функцию учета эффекта произведенного действия и контроля его протекания. Этот факт был установлен наблюдениями, показавшими, что животное без лобных долей мозга оказывается не в состоянии обнаруживать и исправлять допускаемые ошибки, вследствие чего поведение его теряет организованный, осмысленный характер (Прибрам, 1959, 1960, 1961; и др.). Последние годы внесли еще одно существенное дополнение в наше понимание функциональной организации лобных долей мозга животного. Рядом исследователей (Гросс, Вейзкранц, 1964; Брутковский, 1964) было установлено, что лобные доли высших млекопитающих и приматов не являются однородным образованием. Они состоят из нескольких зон. Лобные доли человека развиты неизмеримо больше, чем лобные доли высших обезьян; вот почему у человека процессы программирования, регуляции и контроля сознательной деятельности в несравненно большей степени зависят от префронтальных отделов мозга, чем процессы регуляции поведения у животных. В настоящее время имеется обширный материал, который раскрывает роль префронтальных отделов коры в регуляции психических процессов человека. Отличительная черта процессов регуляции сознательной деятельности у человека заключается в том, что эта регуляция совершается при участии речи (Л. С. Выготский, 1956, 1960; А.Н.Леонтьев, 1959; А.В.Запорожец, 1960; П.Я.Гальперин, 1959). Поэтому есть основания ожидать наиболее грубых нарушений программирующего, регулирующего и контролирующего влияния лобных долей мозга прежде всего на те формы сознательной деятельности, которые совершаются при ближайшем участии речевых процессов. Последние десятилетия принесли нам бесспорные факты, подтверждающие эту гипотезу. Английский исследователь Грей Уолтер (1964, 1966) показал, что ожидание движения на стимул вызывает появление в префронтальной области коры человека медленных потенциалов, которые возрастают по амплитуде по мере увеличения вероятности появления ожидаемого сигнала, уменьшаются с уменьшением этой вероятности и исчезают, как только задача отвечать на сигнал отменяется. Характерно, что эти волны, которые были названы им волнами ожидания, появляются прежде всего в лобных отделах коры и уже оттуда распространяются назад к роландовой борозде. Почти одновременно с этой находкой известный советский физиолог М. Н.Ливанов вместе со своими сотрудниками установили участие префронтальных отделов мозга в наиболее сложных формах активации, вызываемой интеллектуальной деятельностью. Регистрируя с помощью многоканальной установки токи действия, отражающие возбуждение одновременно работающих 100 пунктов мозга, авторы обнаружили, что каждая достаточно сложная умственная работа (например, счет в уме) ведет к увеличению числа синхронно работающих пунктов прежде всего в лобных долях мозга (рис. 5). Р  ис. 5. Изменение корреляции синхронно работающих пунктов лобной области при интеллектуальной деятельности (по М.Н.Ливанову, Н.А.Гавриловой и А. С.Асланову): а — спокойное состояние; б — решение сложной задачи; ис. 5. Изменение корреляции синхронно работающих пунктов лобной области при интеллектуальной деятельности (по М.Н.Ливанову, Н.А.Гавриловой и А. С.Асланову): а — спокойное состояние; б — решение сложной задачи; Таким образом, лобные доли человека принимают самое непосредственное участие в повышении состояния активности, сопровождающем всякую сознательную деятельность. Именно префронтальные отделы коры обеспечивают сложнейшие формы программирования, регуляции и контроля сознательной деятельности человека. Общая физиология сенсорных систем Психические процессы Это одна из групп психических явлений, объединяемых по функциональному назначению. Выделение психических процессов — условное разделение психики на составные элементы. В психических процессах можно выделить два уровня организации: 1 связан с нервными процессами, организуемыми на уровне нейронных связей, данные процессы необязательно выделяются и определяются в сознании личности 2 связан с сознанием и включает в себя познавательные процессы Психические процессы бывают: познавательные, эмоциональные, волевые Первый уровень организации психического процесса – сенсорная система У нее есть периферический, проводниковый и центральный отделы Периферия Рецептор – это специализированная структура (клетка или окончание афферентного нейрона), которая в процессе эволюции приспособилась к восприятию соответствующего раздражителя внутреннего и внешнего мира путем преобразования энергии стимула (раздражителя) в изменение проницаемости своей мембраны. Сенсорные нейроны – биполярные нервные клетки, выполняющие функцию восприятия и проведения возбуждения от периферических рецепторов в ЦНС. Тело находится вне ЦНС, в спинальном ганглии, имеет отросток, который затем Т-образно делится. Один отросток идет на периферию и образует там чувствительные окончания (рецепторы). Другой отросток идет в ЦНС, где ветвится и формирует синаптические окончания. Виды рецепторов Хеморецепторы — рецепторы, чувствительные к воздействию химических веществ. Белковый комплекс, который, взаимодействуя с определённым веществом, изменяет свои свойства, что вызывает каскад внутренних реакций организма. Среди таких рецепторов: рецепторы органов чувств (обонятельные и вкусовые рецепторы) и рецепторы внутреннего состояния организма (рецепторы углекислого газа дыхательного центра, рецепторы рН внутренних жидкостей). Механорецепторы — это окончания чувствительных нервных волокон, реагирующие на механическое давление или деформацию. Среди них: тельца Мейснера, тельца Меркеля, тельца Руффини, тельца Пачини, мышечные веретена, сухожильные органы Гольджи, рецепторы вестибулярного аппарата. Ноцицепторы — болевые рецепторы. Расположены главным образом в коже (кожные ноцирецепторы) или во внутренних органах (висцеральные ноцирецепторы). В окончаниях миелинизированных волокон (А-тип) они реагируют только на механическое раздражение; в окончаниях немиелинизированных волокон (С-тип) могут реагировать на различные типы раздражений (механическое, тепловое или химическое). Фоторецепторы — светочувствительные нейроны. Содержатся во внешнем зернистом слое сетчатки. Терморецепторы — отвечают за температурную рецепцию. Основные из них: колбочки Краузе (ощущение холода) и тельца Руффини (ощущение тепла). Структурно-функциональная организация рецепторов: – первичночувствующие – обоняние, тактильные, проприорецепторы (восприятие стимула осуществляется непосредственно окончанием афферентного нейрона); – вторичночувствующие: вкус, слух, зрение, вестибулорецепторы (здесь между действующим стимулом и афферентным нейроном располагается специализированная клетка эпителиального происхождения, из которой при раздражении выделяется медиатор, действующий на окончание афферентного нейрона). Чувствительность включает в себя 3 основных этапа: 1. взаимодействие стимула с белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки; 2. внутриклеточные процессы усиления и передачи сенсорного стимула; 3. открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникает рецепторный потенциал). В первичночувствующих рецепторах рецепторный потенциал является одновременно и генераторным. У вторичночувствующих – рецепторный потенциал вызывает выделение квантов медиатора из пресинаптических окончаний рецепторной клетки. Медиатор изменяет поляризацию ПСМ. Общие принципы строения сенсорных систем многоканальность, т.е. наличие в каждом слое множества нервных клеток, связанных с множеством клеток следующего слоя. Параллельные каналы передачи и обработки информации обеспечивают точность, детальность анализа сигналов и большую надежность. Дублирование необходимо для повышения надёжности системы. Возбуждение передается от рецепторов в кору всегда по нескольким параллельным путям. Потоки возбуждения частично дублируются, и частично разделяются. По ним передается информация о различных свойствах раздражителя. Пример параллельных путей зрительной системы: 1-й путь: сетчатка — таламус - зрительная кора. 2-й путь: сетчатка - четверохолмие (верхние холмы) среднего мозга (ядра глазодвигательных нервов). 3-й путь: сетчатка — таламус - подушка таламуса - теменная ассоциативная кора. При повреждении разных путей и результаты получаются различные. Например : если разрушить наружное коленчатое тело таламуса (НКТ) в зрительном пути 1, то наступает полная слепота; если разрушить верхнее двухолмие среднего мозга в пути 2, то нарушается восприятие движения предметов в поле зрения; если разрушить подушку таламуса в пути 3, то пропадает узнавание предметов и зрительное запоминание. Во всех сенсорных системах обязательно существуют три пути (канала) передачи возбуждения: 1) специфический путь: он ведет в первичную сенсорную проекционную зону коры, 2) неспецифический путь: он обеспечивает общую активность и тонус коркового отдела анализатора, 3) ассоциативный путь: он определяет биологическую значимость раздражителя и управляет вниманием. В эволюционном процессе усиливается многоэтажность и многоканальность в структуре сенсорных путей. 2. многослойность (многоэтажность). В каждой сенсорной системе существует несколько передаточных промежуточных инстанций на пути от рецептора к коре ГМ. В каждом слое происходит частичная специализированная переработка информации. Это дает возможность организму быстро реагировать на простые сигналы, анализируемые уже на первых уровнях, а также избирательно регулировать свойства нейронных слоев под влиянием из других отделов мозга. Например: Мошка летит прямо в глаз - глаз моргнул в ответ, и мошка в него не попала. Для ответной реакции в виде моргания не требуется создавать полноценный образ мошки, достаточно простой детекции того, что объект быстро приближается к глазу. Одна из вершин многоэтажного устройства сенсорной системы - это слуховая сенсорная система. В ней можно насчитать 6 этажей. Существуют также дополнительные обходные пути к высшим корковым структурам, которые минуют несколько низших этажей. Таким способом кора получает предварительный сигнал для повышения её готовности до основного потока сенсорного возбуждения. 3. Принцип конвергенции. Конвергенция — это схождение нервных путей в виде воронки. За счёт конвергенции нейрон верхнего уровня получает возбуждение от нескольких нейронов нижележащего уровня. Например: в сетчатке глаза существует большая конвергенция. Фоторецепторов несколько десятков млн., а ганглиозных клеток - не более одного млн. Т.е. нервных волокон, передающих возбуждение от сетчатки во много раз меньше, чем фоторецепторов. Воронка «Шеррингтона». Физиологический смысл «суживающейся воронки» — в уменьшении избыточности информации. Принцип общего конечного пути по Чарльзу Шеррингтону. В литературе иногда используется термин «воронка Шеррингтона». «Согласно его представлениям, количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути. Благодаря такому столкновению достигается блокирование всех воздействий, кроме одного, которое и регулирует протекание рефлекторной реакции. Для пояснения этого принципа часто используют метафору: предположим, на железнодорожную станцию по пяти путям прибывает пять составов, но со станции отходит только один путь и, соответственно, со станции в единицу времени уйдёт только один поезд. Таким образом, уже сами принципы организации нервной системы предполагают, что только некоторые из внешних воздействий в условиях их одновременного влияния на организм получат на выходе «доступ» к мышцам. Сам Чарльз Шеррингтон считал, что важнейшим фактором, обеспечивающим выбор одного из нескольких возможных воздействий, является сила воздействия: сильное воздействие как бы подавляет, вытесняет более слабые. 4. Принцип дивергенции. Дивергенция - это расхождение потока возбуждения на несколько потоков от низшего этажа к высшему (напоминает расходящуюся воронку). Физиологический смысл – в обеспечении дробного и сложного анализа разных признаков сигнала. 5. Принцип обратной связи. Обратная связь обычно означает влияние управляемого элемента на управляющий. Для этого существуют соответствующие пути возбуждения от низших и высших центров обратно к рецепторам. 6. Принцип кортикализации (в новой коре представлены все сенсорные системы; следовательно, кора функционально многозначна, и не существует абсолютной локализации) 7. Принцип двусторонней симметрии (существует в относительной степени) 8. Принцип структурно-функциональных корреляций Любая психофизиологическая функция зависит от одновременной работы нескольких сенсорных систем, то есть является полисенсорной, и поэтому не может быть локализованной в ограниченных отделах мозга. Сама же сенсорная система выступает как определенная локализованная анатомическая система, выполняющая специализированную функцию обнаружения и преобразования информации в нервный код, в котором заключена совокупность описания признаков воспринимаемого объекта или явления. 9. дифференциация по вертикали и горизонтали. По вертикали – образование отделов, состоящих из нескольких нейронных слоев и осуществляющих определенную функцию. По горизонтали различные свойства рецепторов, нейронов и связей между ними в пределах каждого из слоев. |