колок по опорно двиг и покровам клетки. Опорнодвигательная система клетки
 Скачать 2.8 Mb. Скачать 2.8 Mb.
|

|
Коллоквиум № 4. "Опорно-двигательная система клетки". Основные вопросы: Дать сравнительную характеристику молекулярно-структурной организации промежуточных филаментов, микрофиламентов и микротрубочек. Назвать основные белки и параметры их агрегации.
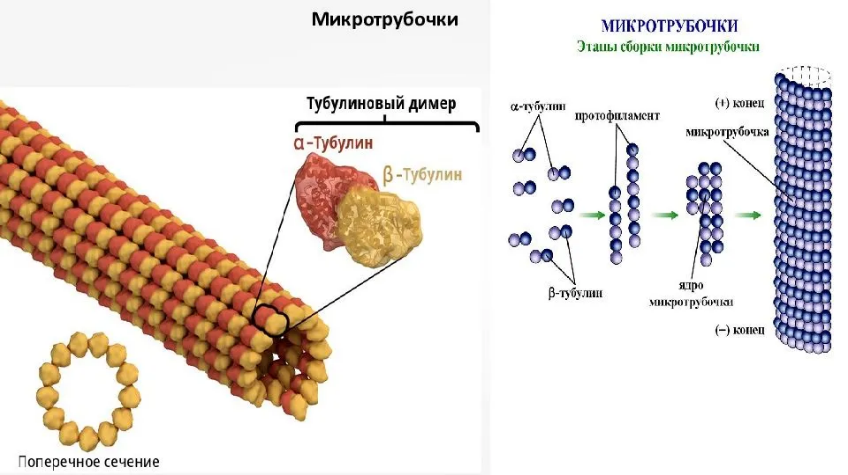
Для каких цитоскелетных структур характерны процессы самосборки и разборки: Что известно об этих механизмах? Микрофиламенты способны к самосборке только при высокой концентрации актина в сдвоеном спиральную ленту толщино 6-7 нм (эта актиновая фибрила обладает полярность - + и – конец. Со стороны + конца происходит полимеризация (растёт), а со стороны – конца происходит деполимеризация (убывает). Полимеризация актина носит обратимый характер. Благодаря изменению концентрации G- и F-форм актина изменяется и вязкость цитоплазмы.  Микротрубочки также способны к самосборке Процесс самосборки происходит в центрах организации микротрубочек(ЦОМТ) (локализованный в клеточном центре). В клеточном центре лежат 2 центриоли, от них отходят во все направления микротрубочки, образующие радиальный скелет и поддерживая её форму. В качестве центра организации микротрубочек (ЦОМТ) на центриолях функционирует особые булавовидные образования – сателлиты и тяготящие к ним плотные тельца – фокусы схождения микротрубочек. В зонах ЦОМТ осуществляется сборка микротрубочек, обращенных к клеточному центру – концом (заблокирована, чтобы микротрубочка не развалилась), + конец обращен в мембрану. Микрофиламенты как скелетные и двигательные структуры. При каких условиях возникает способность микрофиламентов к двигательной реакции? Объясните механохимические основы мышечного движения. Скелетная: под плазматической мембраной микрофиламенты часто образуют густую сеть – кортикальный скелет. Стабилизация формы клетки. Также амебоидное движение за счёт разжижжения цитоплазмы. Также мембрану можно выгибать благодаря надстраивающейся системе микрофиламентов – образуя филоподии и ламелоподии. Двигательная: для движения используется белок миозин. Миозин состоит из 2х головок и 2х переплетённых хвостиков. Основная особенность миозина является способность расщеплять АТФ и за счёт этого изменяет свою конформацию, при этом головка миозина наклоняетсяпо отношению к хвостику, меняя свой угол наклона. Дать сравнительную характеристику организации акто-миозиновых комплексов в немышечных (фибробласт, амеба) и специализированных мышечных клетках (поперечно-полосатое мышечное волокно). Показать иммуноцитохимическую локализацию альфа-актинина, актина и миозина в этих структурах. Моизин топает по актину  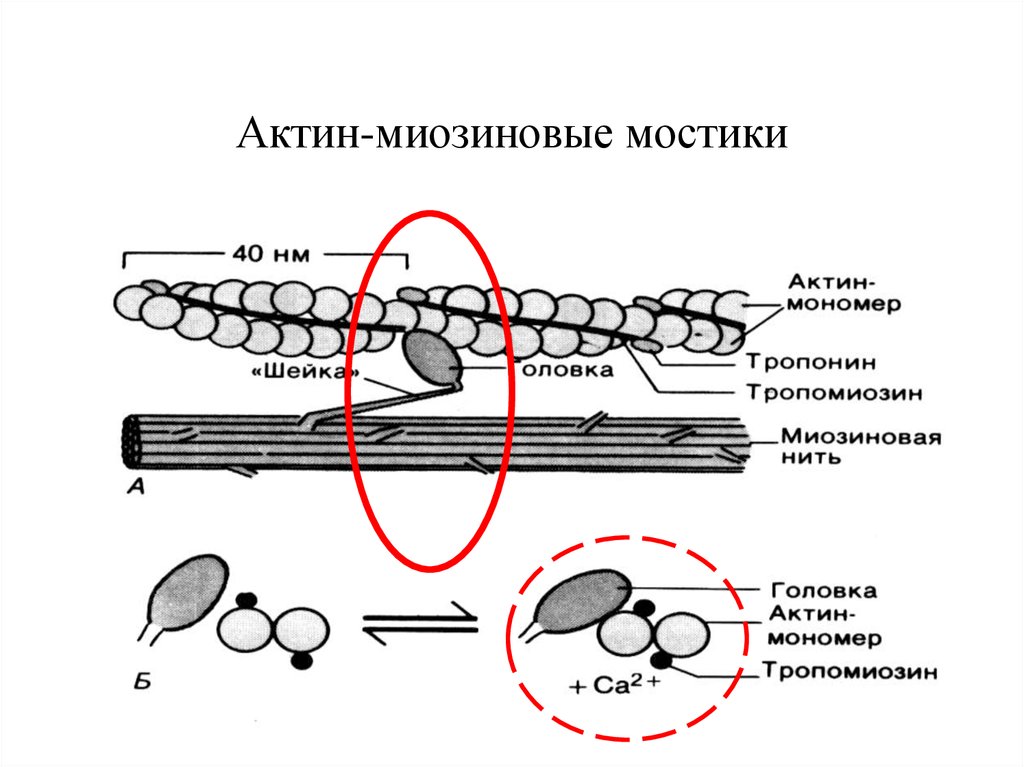 Строение саркомера:  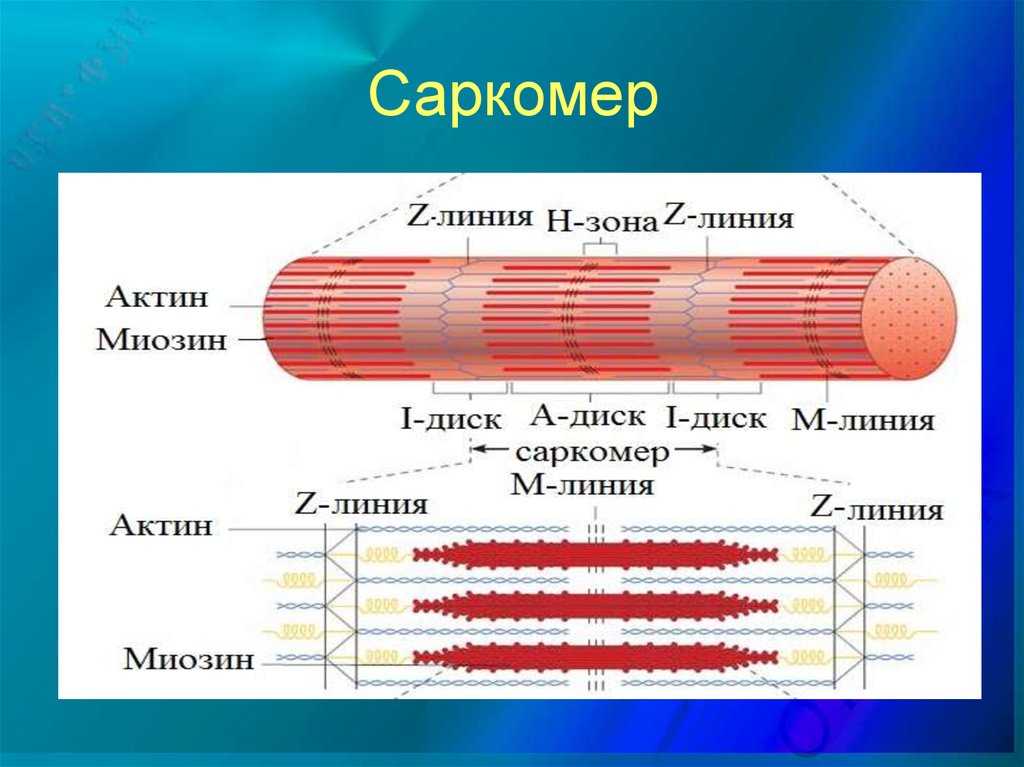 В поперечнополосатых мышечных волокнах миофибриллы имеют периодическую организацию: актиновые филаменты и пучки миозина собраны в правильные блоки – саркомеры. На стыках саркомеров (в Z-полосках) микрофиламенты скреплены α-актинином. В соседних миофибриллах саркомеры и их диски лежат на одном уровне, вследствие чего всё мышечное волокно представляется поперечно исчерченным – «поперечнополосатым» Строение актино-миозинового комплекса одноклеточных 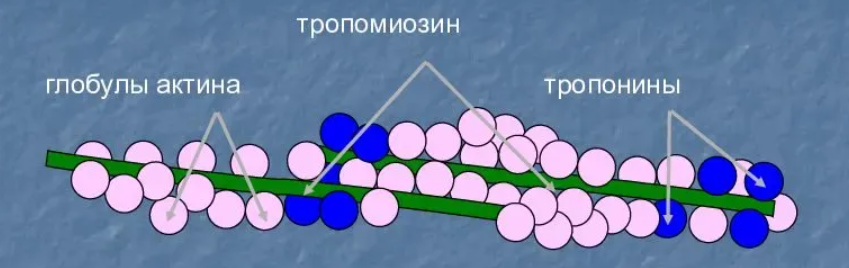 При движении клетки по субстрату в ламеллоподиях ведущего (переднего) края кортикальная сеть филаментов разбирается, цитоплазма разжижается и натекает на плазмалемму. «Натёк» стабилизируется восстановлением кортикальной сети филаментов. При этом клетка крепится к субстрату при помощи специальных фокальных контактов, возникающих впереди и разрушающихся сзади. Механическое натяжение создается пучками актиновых филаментов – стресс-фибриллами, закрепленными на фокальные контакты с помощью белка альфа-актинина. Такая сеть пронизывает весь объем клетки. При включении в сеть белка миозина возникают сократимые актомиозиновые комплексы. Тело клетки передвигается от заднего конца к переднему. 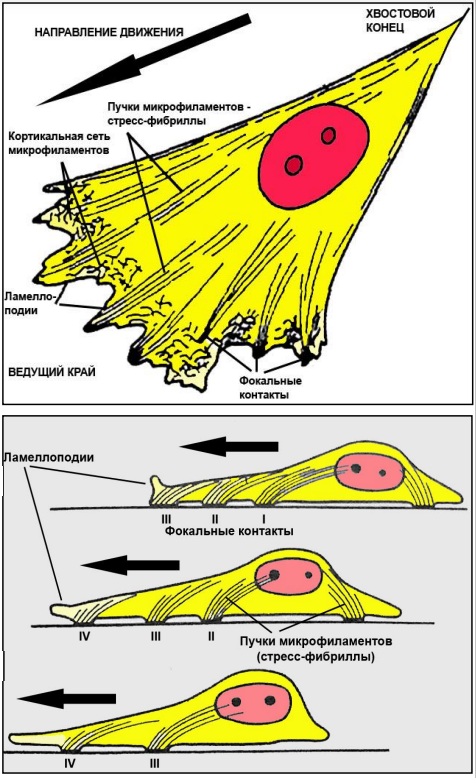
В МЫШЕЧНОЙ ТАКЖЕ КАК И В НЕ МЫШЕЧНОЙ ПРОСТО КОМПЛЕКС МЕНЬШЕ Микротрубочки как скелетные и двигательные структуры. Привести примеры использования микротрубочек в качестве цитоскелета. Полярная молекула состоящая из глобулярного белка тубулин – альфа- и бета- Скелетная: Стабилизация формы клетки, создавая её радиальный скелет – астросферу. Двигательная: осуществляется за счёт двигательных белков (динеин и кинезин), которые способны перемещаться по тяжам тубулина «шажками» при этом изменяя свою конформацию, они присоединяются одним концом к органоиду/не мембранным внутриклеточным структурам. NB: Сами центриоли к росту цитоплазматических микротрубочек отношения не имеют. Центриоли – это резервированные базальные тельца киноцилий – ресничек и жгутиков. Они каким-то образом по своему подобию программируют и направляют сборку микротрубочек осевого стержня киноцилии – аксонемы – по схеме «9х2 + 2» Показать строение реснички и объяснить механохимические основы ресничного движения. Строение ресничек 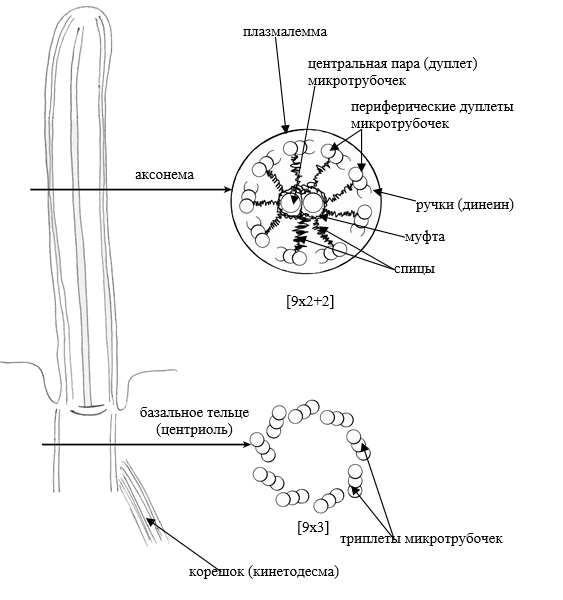 •Механизм движения киноцилий связан с активностью моторного белка динеина. Молекулы динеина образуют «ручки», которыми одна пара микротрубочек взаимодействует с другой парой. На правой половине аксонемы динеиновые ручки «проползают» по поверхности микротрубочек (с расщеплением АТФ, т.е. с участием энергии) и вызывают продольное смещение соседних пар микротрубочек. На левой половине аксонемы динеиновые ручки расслаблены, не работают. Этого достаточно для изгибания реснички (жгутика) в правую сторону. 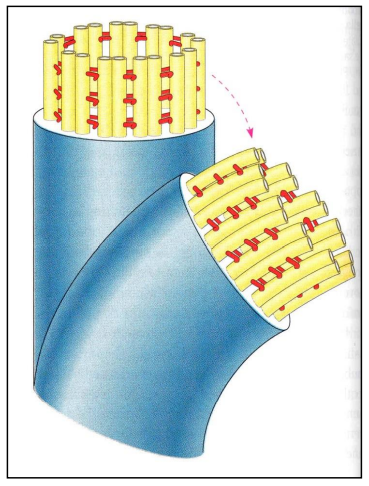 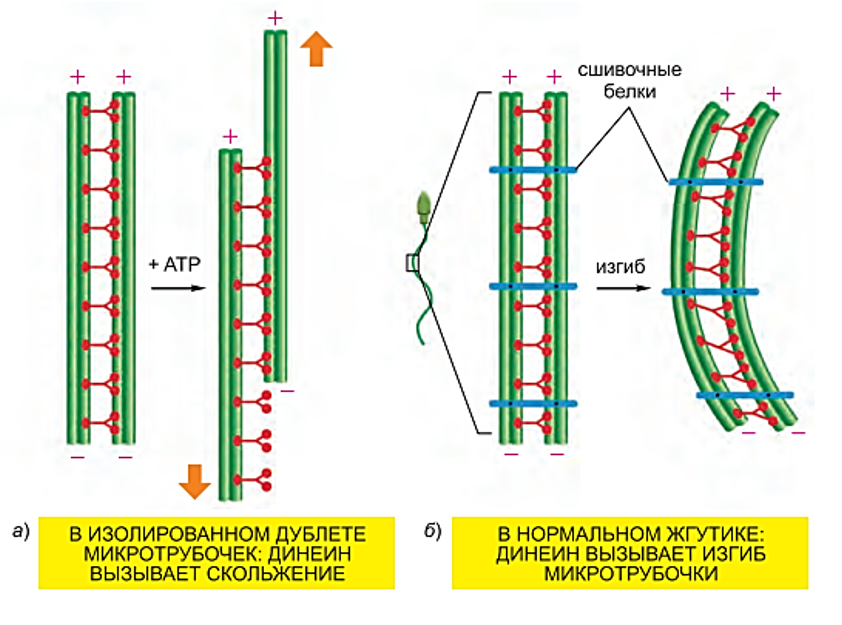 Что является центрами организации микротрубочек в клетке? Где они локализуются? Центр Организации микротрубочек (ЦОМТ) – организованные участки клетки, в которых происходит полимеризация микротрубочек. Локализован в клеточном центре выполняют две основные функции: организацию эукариотических жгутиков и ресничек и организацию митотического и мейотического веретенообразного аппарата,которые разделяют хромосомы при делении клеток. ЦОМТ является основным местом зарождения микротрубочек и может быть визуализирован в клетках путем иммуногистохимического обнаружения γ-тубулина. Морфологические характеристики ЦОМТ варьируются между различными типами и царствами. [1] У животных двумя наиболее важными типами ЦОМТ являются: 1) базальные тела, связанные с ресничками и жгутиками, и 2) центросомы, связанные с образованием веретена. Строение центриолей, их взаиморасположение в клеточном центре. Механизм воспроизведения центриолей. В основе клеточного центра лежат 2 центриоли, эта пара называется диплосома/центросома. Диплосома состоит из 2х центриолей: материнской и дочерней. Центриоли образованы девяти триплетами микротрубочек. В качестве ЦОМТ на центриолях функционируют особые белковидные образования – сателлиты – и тяготящие к ним плотные тельца – фокусы схождения микротрубочек, стягивающие на себя пучки микротрубочек. . В зонах ЦОМТ осуществляется сборка микротрубочек, обращенных к клеточному центру – концом (заблокирована, чтобы микротрубочка не развалилась), + конец обращен в мембрану. Когда центриоли расходятся к полюсам делящейся клетки, бывшая дочерняя центриоль подвергается перестройке и приобретает сателлиты, – теперь она тоже может выступать в качестве клеточного центра. Микротрубочки, отрастающие теперь от обеих центриолей, направлены от полюсов к экватору и играют роль веретена деления. По микротрубочкам веретена с помощью особой формы динеина будут перемещаться хромосомы – от центра к полюсам клетки. У ценриолей в клетке имеется еще одна важная функция – именно они выступают в качестве базальных телец ресничек и жгутиков (см. выше). Следует заметить, что центриоли отсутствуют в клетках голосеменных и цветковых растений. Это связано с тем, что у их 74 клеток жгутики или реснички не обнаруживаются ни на одной стадии жизненного цикла, в отличие от всех остальных высших растений, включая мхи и папоротники. 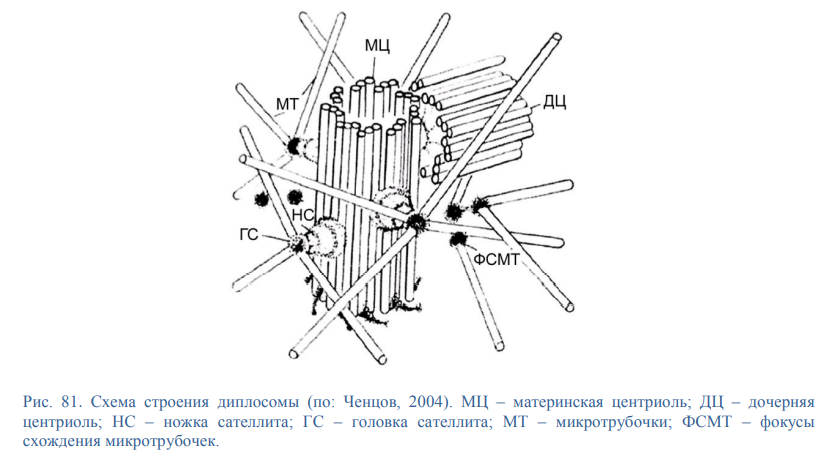 Какова роль центриолей и как объяснить их отсутствие в клетках высших растений? См Выше Дополнительные вопросы: Провести аналогию в механохимической организации и принципах работы актин-миозиновых и тубулин-динеиновых двигательных систем. Какие опорные и двигательные структуры имеются у растительных клеток? Как они развиты по сравнению с животными клетками? Как организован двигательный аппарат бактерий? Каковы принципиальные отличия бактериального жгутика от реснички эукариотной клетки? Коллоквиум № 5. "Поверхностный аппарат клетки". Основные вопросы: Назвать составные части поверхностного аппарата клетки, определить их функции. Выделяют три составные части поверхностногоаппарата: плазматическую мембрану (плазмалемму) – липо-протеиновый пленочный комплекс, в состав которого входят липиды (фосфолипиды), формирующий билипидный слой, в котором для осуществления транспорты находятся белки ответсвенные з атранспорт различных веществ Ф-ция плазмалеммы: защита от патогенов и воздействий окр. среды, транспорт необходимых для клетки веществ надмембранный комплекс (гликокаликс) – совокупность разветвленных гликопротеидов и гликолипидов на поверхности мембраны. Ф-ция: специфична для клеток разных тканей. Микроворсинки кишечника – сорбция ферментов, пристеночное пищеварение, всасывание Осморегулируещие и выделительные эпителии – создает локальное повышение концентрации ионов для их всасывания. Влияет на состояние околоклеточной среды – создание отрицательного заряда на поверхности эритроцитов для предотвращения их агглютинации (склеивание с антигенами) Рецепторно-сигнальная ф-ция Обеспечивает адгезию (избирательное сцепление клеток и другиз частей) субмембранный комплекс (кортикальный цитоскелет) - Химический состав и организация плазматической мембраны. Привести доказательства в пользу жидкостно-мозаичной модели. Липидный и белковый состав различается в мембранах разных органоидов, а также в разных участках плазматической мембраны, в том числе, с ее двух сторон. Это соотношение определяется функциями той или иной мембраны. Строение мембранных белков таково, что с водной фазой они соприкасаются гидрофильными аминокислотами, а с липидной фазой – гидрофобными, что стабилизирует их положение в мембране. В свою очередь, липидная фаза мембраны стабилизируется белками, имеющими связь с субмембранным цитоскелетом, так что белковый каркас не дает липидам мембраны растечься. Такая организация плазматической мембраны определяется как жидкостно-мозаичная модель; она подразумевает, что в жидком слое постоянно мигрирующих липидов в мозаичном порядке встроены белки, заякоренные на белки цитоскелета и потому гораздо менее подвижные. Таким образом, положение белков в мембране контролируется клеткой. Благодаря связыванию липидов с белками мембрана ведѐт себя как жидкостное образование, но при этом остаѐтся стабильной. Несмотря на то, что на место одних липидов постоянно встают другие, общая мозаика липидов и белков не изменяется. Гликокаликс: химический состав, степень развития, свойства и функции в специализированных клетках. Надмембранные структуры клеток растений, грибов, бактерий.  В состав гликокаликса, помимо углеводов, могут входить и белки. Примером таких белков служат специфические рецепторы В-лимфоцитов – иммуноглобулины. Основная часть молекулы иммуноглобулина закреплена в мембране с помощью одного гидрофобного участка, а рабочая, вариабельная часть, несущая антигенные детерминанты, находится в надмембранной области. Еще одной разновидностью надмембранного комплекса являются полисахаридные клеточные стенки бактерий, грибов и растений – целлюлозные, муреиновые, хитиновые и др. У некоторых животных развитие надмембранных структур в клетках кожного эпителия приводит к образованию кутикул, например, хитиновых оболочек у членистоногих и тунициновых – у асцидий. В состав гликокаликса, помимо углеводов, могут входить и белки. Примером таких белков служат специфические рецепторы В-лимфоцитов – иммуноглобулины. Основная часть молекулы иммуноглобулина закреплена в мембране с помощью одного гидрофобного участка, а рабочая, вариабельная часть, несущая антигенные детерминанты, находится в надмембранной области. Еще одной разновидностью надмембранного комплекса являются полисахаридные клеточные стенки бактерий, грибов и растений – целлюлозные, муреиновые, хитиновые и др. У некоторых животных развитие надмембранных структур в клетках кожного эпителия приводит к образованию кутикул, например, хитиновых оболочек у членистоногих и тунициновых – у асцидий.Гипертрофия гликокаликса приводит к образованию клеточных оболочек (стенок) у бактерий, грибов, растений или кутикул у ряда животных. Свойства: создание околоклеточной среды, способен запутывать в своих полисахаридных ветвях ионы, способность придавать клеткам общий хим. состав благодаря чего они и узнаю друг друга, благодаря чрезвычайному разнообразую химических связей в молекулах углеводов, последние являются специфичными маркерами, Ф-ция: рецепторная, околоклеточной среды, роль ионных «ловушек», пристеночное пищеварение (с участием ферментов), отчасти тканеспецифичная узнаваемость клеток благодаря маркерам, образуют клеточные стенки. Кортикальный (субмембранный) цитоскелет: его элементы, связь с плазмалеммой. Механизмы и значение латерального перемещения белков плазмалеммы. Субмембранный комплекс образован элементами периферического примембранного цитоскелета (кортикального цитоскелета) и белками, обеспечивающими его связь с мембраной. Занимает промежуточное положение между гиалоплазмой и плазмалеммой Взаимодействие кортикальных актиновых филаментов с миозином и их скольжение друг относительно друга составляют механизм движения микроворсинок в клетках кишечного эпителия, образования перетяжки при клеточном делении, сужения апикальных (верхних) концов клеток эктодермального пласта при формировании нервной трубки в эмбриогенезе и других процессов. Важную функцию выполняет периферический цитоскелет в яйцеклетках: в его сети, в строго определенных местах зафиксированы особые регуляторные молекулы – морфогенетические факторы, которые в последствии, при делении зиготы, попадут в разные клетки зародыша и предопределят различные пути развития этих клеток. Функционирование кортикального цитоскелета во многом определяет подвижность клеточной поверхности и ее отдельных белков. Взаимодействие плазмалеммы с подвижным цитоскелетом (микрофиламентами и микротрубочками) позволяет перемещать мембранные белки, рецепторы по поверхности клетки для поглощения веществ (эндоцитоз) или для сбрасывания «мусора» Обновление и рост плазматической мембраны. Какие субмембранные и цитоплазматические структуры задействованы в этих процессах? Как оценивать скорость обновления и роста плазмалеммы? Рост и обновление осуществляется за счёт везикулов аппарата Гольджи, в процессе компенсаторного пиноцитоза и выноса новых мембранн аппарта Гольджи. Связь плазмалеммы с цитоскелетом и ее текучесть обеспечивают возможность отрыва и переноса микровезикул. Скорость роста – удвоение площади за 1 мин. Механизм роста – продукция пузырьков аппаратом Гольджи и их интеркаляция (встраивание) в мембрану (экзоцитоз) Охарактеризовать формы трансмембранного транспорта веществ: диффузию, пассивный и активный транспорт. С какими элементами плазмалеммы связаны эти формы транспорта? Диффузия: Если концентрация солей во внешней среде выше, чем внутри клетки, то вода будет диффундировать через мембрану во внешнюю среду, разбавляя концентрированный раствор до тех пор, пока концентрация соли по обе стороны мембраны не уравняется. Происходит через билипидный слой Пассивный транспорт(облегчённая диффузия)– транспорт, не требующий затраты энергии, происходит по градиенту концентрации. Для того чтобы проходила не только вода, но и ионы в мембране находятся специальные белки. Через мембранные поры Активный транспорт – транспорт, требующий затрат энергии, происходит против градиента концентрации. Белки переносчики, либо белки, которорые обратимо формируют свою конфротацию и пропускают молекулы. Отчего возникает концентрационный гетероградиент ионов и как он используется клеткой? Гетероградиент ионов – разница в концентрации отдельных ионов с двух сторон от плазмалеммы (с внутренней стороны клетки и с внешней стороны); жизненно необходим клеткам для облегчения проникновения веществ при пассивном транспорте; используется также для проведения электрического возбуждения вдоль мембраны, в частности, нервных клеток. Один из важнейших насосов – калий-натриевый (K+/Na+) насос. Этот антипортный канал за один цикл, используя 1 молекулу АТФ, выкачивает из клетки 3 иона Na+ и закачивает 2 иона K+. ● В результате на мембране возникает разность электрических потенциалов с положительным зарядом снаружи (преобладание Na+) и отрицательным изнутри (преобладание анионов над катионами). ● Электропотенциал – универсальное для всех живых клеток свойство: пока клетка жива, её плазмалемма электрополяризована.(см. картинку) ● В результате действия различных насосов, локализованных в плазмалемме (для Н+ , Na+ , K+ , Ca2+, Cl- и др.), с учетом перетока многих ионов в ЭПР и их связывания с белками, разность потенциалов между наружной поверхностью и цитоплазмой в разных клетках составляет от +15 до +80 мВ (милливольт). ● Градиент концентрации катионов (Na+ , H+ ), создаваемый за счет работы насосов, используется во всех клетках для питания. ● Вместе с ионами натрия или протонами через каналы пассивного транспорта в клетку поступают глюкоза, аминокислоты и другие пищевые мономеры. За пределами клетки мономеры связываются с катионами, чтобы использовать их как «пропуск» для прохождения через ионные каналы (симпорт), а затем, попадая в цитоплазму, отсоединяются. ● Таким образом, поддержание градиента концентрации ионов необходимо для транспорта питательных веществ и является неотъемлемой частью клеточного гомеостаза. ● На поддержание своего гомеостаза клетка тратит 80 % всей АТФ! ● Особое значение разность зарядов приобретает в нервных клетках при создании нервного импульса. В момент возбуждения нейрона внешним раздражителем (химическим или физическим) происходит кратковременное раскрытие каналов пассивного транспорта, и скопившиеся на внешней стороне мембраны ионы Na+ вбрасываются в клетку по градиенту концентрации. В результате осуществляется кратковременная (0,001 секунды) локальная деполяризация мембраны нейрона и распространение электромагнитной волны по мембранам отростков, что и составляет сущность передачи нервного импульса. Что такое микроворсинки? У каких клеток они развиты, как устроены и как работают? 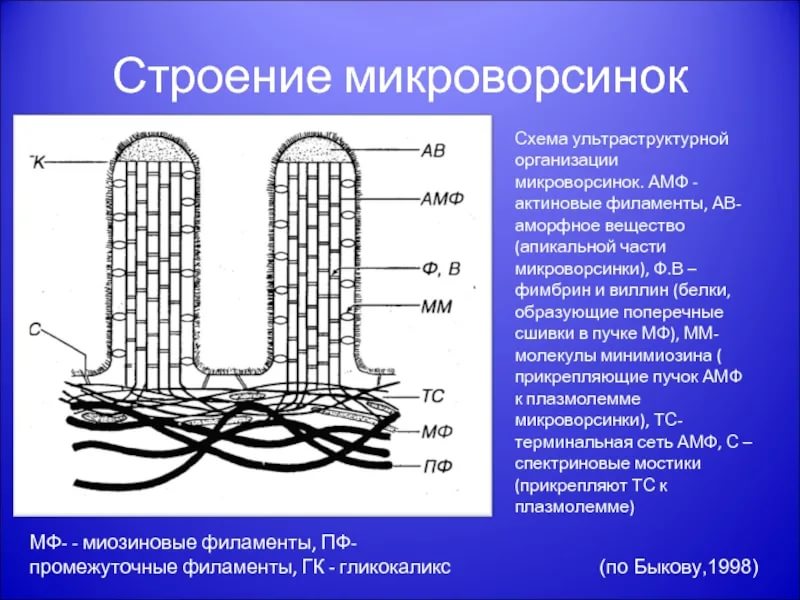 Микроворсинками называют пальцевидные выросты плазматической мембраны некоторых животных клеток. Иногда микроворсинки увеличивают площадь поверхности клетки в 25 раз, поэтому они особенно многочисленны на поверхности клеток всасывающего типа, а именно в эпителии тонкого кишечника и извитых канальцев нефронов. Это увеличение площади всасывающей поверхности способствует и лучшему перевариванию пищи в кишечнике, потому что некоторые пищеварительные ферменты находятся на поверхности клеток и связаны с ней. Микроворсинками называют пальцевидные выросты плазматической мембраны некоторых животных клеток. Иногда микроворсинки увеличивают площадь поверхности клетки в 25 раз, поэтому они особенно многочисленны на поверхности клеток всасывающего типа, а именно в эпителии тонкого кишечника и извитых канальцев нефронов. Это увеличение площади всасывающей поверхности способствует и лучшему перевариванию пищи в кишечнике, потому что некоторые пищеварительные ферменты находятся на поверхности клеток и связаны с ней.Охарактеризовать формы и механизмы транспорта веществ в мембранной упаковке. Привести примеры использования эндоцитоза и экзоцитоза в жизнедеятельности тканевых клеток и одноклеточных организмов. Макромолекулы и более крупные частицы транспортируются в клетку посредством везикулярного транспорта, т. е. будучи заключенными в мембранную оболочку ● Везикулярный транспорт возможен в двух направлениях: экзоцитоз – вынос веществ из клетки (секреция, экскреция); эндоцитоз – поглощение веществ. ● Эндоцитоз в свою очередь может протекать как фагоцитоз – поглощение крупных частиц, и пиноцитоз – поглощение водных растворов с помощью мельчайших везикул. ● При этом активно используются гликокаликс – распознавание «пищи», и субмембранный цитоскелет – втягивание и выталкивание мембран. ● Пиноцитоз и фагоцитоз – ключевые реакции жизнедеятельности ряда специализированных клеток. ● Они составляют основу пищевого поведения амебоидных протистов, а также основу функционирования фагоцитарных клеток многоклеточного организма, таких как макрофаги и нейтрофилы. Что такое клеточные рецепторы? Какова их локализация и химическая природа? Клеточный рецептор — молекула (обычно белок или гликопротеин) на поверхности клетки, клеточных органелл или растворенная в цитоплазме. ● Специфично реагирует изменением своей пространственной конфигурации на присоединение к ней молекулы определённого химического вещества, передающего внешний регуляторный сигнал и, в свою очередь, передает этот сигнал внутрь клетки или клеточной органеллы, нередко при помощи так называемых вторичных посредников или трансмембранных ионных токов. ● Рецепция – способность клетки принимать внешние сигналы и передавать соответствующую информацию на внутриклеточные структуры для формирования ответной реакцией. ● Разнообразие сигналов: - физические (свет, рельеф), химические (веществалиганды), биологические (вирусы, чужие клетки). ● Разнообразие ответов: - цитотаксисы (направленное движение), выброс секретов, мышечное сокращение, клеточное деление и дифференцировка, иммунный ответ и др. ● Рецепторы – это обычно трансмембранные гликопротеиды с преобладающим наружным доменом; редко гликолипиды. ● Эффекторы – обращенные в цитоплазму белки, формирующие внутриклеточные сигналы для генов и рабочих структур клетки. ● Общие свойства рецепторов. - Специфичная молекулярная конфигурация рецепторных молекул обеспечивает их высокую избирательность к различным сигнальным лигандам – гормонам и др. БАВ, а также тканеспецифичный сигналинг. ● Число рецепторов в плазмалемме относительно невелико (500-100000 рецепторов на клетку, это менее 0,1 % всех белков мембраны). – ● Связывание лигандов кратковременно и обратимо. ● Механизм рецепции связан с кратковременным изменением конформации белка-рецептора, которое генерирует новый внутриклеточный сигнал. ● По типу эффектора (по механизму формирования внутриклеточного сигнала) различаются рецепторы: каналообразующие (ионофорные) и каталитические (ферментные). Объяснить два способа инициации внутриклеточного сигнала: аденилатциклазный и ионофорный. Сравнить их на примере рецепторов пептидных гормонов (глюкагона и инсулина) и нейромедиатора (ацетилхолина). Каналообразующие рецепторы ● В каналообразующем рецепторе эффектором является ионный канал – ионофор ● При раздражении рецептора канал раскрывается, и происходит быстрый вброс ионов Na+ , кратковременная (0,001 сек) деполяризация мембраны и её электромагнитное возбуждение. ● Ответ – физиологическое возбуждение клетки. ● Фоторецепторы(см.картинку) Нейромышечный синапс: нейромедиатор ацетилхолин выбрасывается из нервного окончания и связывается с рецептором на плазмалемме мышечного волокна. Возбужденный рецептор активирует (раскрывает) ионные Na+ -каналы. Ответ – сокращение миофибрилл. ● Каталитические рецепторы имеют другие эффекторы – примембранные ферменты, производящие различные сигнальные молекулы – вторичные мессенджеры. ● Аденилатциклаза – производит циклический АМФ (цАМФ) – активатор многих сигнальных путей; ● Фосфолипаза С – дает другие мессенджеры, в т.ч. Ca2+ . ● G-белок – ГТФ связывающий ±регулятор эффекторов. ● При раздражении рецептора лигандом с этих ферментов начинается каскад реакций вторичных мессенджеров. В основном это фосфорилирующие ферменты сигнальных путей клетки – протеинкиназы. Сигнал проходит на рабочие структуры клетки, вплоть до ядра, активируя одни гены и подавляя другие. В итоге регулируются синтезы разнообразных белков, деление, дифференцировка и рост клеток. ● Сигнальный путь растительного гормона роста ауксина – от рецептора до генов и цитоплазматических структур. (см.картинку) ● Аденилатциклазная система регуляции может объединять несколько гормонов и рецепторов. Их влияние на сигнальный путь опосредуется через G-белок, который может активировать или, напротив, ингибировать эффектор (аденилатциклазу). ● Вторичные мессенджеры: цАМФ (cAMP) и каскад протеинкиназ, фосфорилирующих белки сигнальных путей. ● При взаимодействии 2 рецепторов создаются гибкие и устойчивые системы саморегуляции по принципу обратной отрицательной связи. ● Пример: регуляция поглощения и выделения глюкозы гепатоцитами. Взаимодействуют 2 сигнальных пути: 1. Глюкагон – рецептор – G-белок (+) – активация аденилатциклазы – цАМФ – протеинкиназы – расщепление гликогена до глюкозы – вынос глюкозы из клетки. 2. Инсулин – его рецептор – раскрытие глюкозо-Na+ канала (вход глюкозы) и одновременное действие на G-белок (-) – блокировка аденилатциклазы и прекращение распада гликогена. Перечислить и показать локализацию постоянных межклеточных контактов в кишечном эпителии. В чем особенность химических (транспортных) контактов и где они встречаются? ● Адгезия – прилипание клеточной поверхности к субстрату или другим клеткам с помощью клейкого гликокаликса. ● Простая адгезия типична для клеток эмбриональных листков, зачатков, где постоянные контакты еще не сформированы. ● Адгезия усиливается замками типа «шип-паз». Адгезия между клетками или на межклеточный субстрат (матрикс) происходит с участием особых трансмембранных адгезивных гликопротеидов (CAM – cell adhesion molecules), включенных в гликокаликс (кадгерины, селектины, интегрины и др.). ● Включение Ca2+ усиливает сцепление. Адгезия контролируется рецепторами плазмалеммы, поэтому она ткане- и видоспецифична (опыты по реагрегации эмбрионов, губок). Специальные межклеточные контакты ● Более сложные контакты образуются в тканях. ● У животных различаются следующие виды клеточных контактов: заякоревающие (сцепляюшие), запирающие (плотные), щелевые и синаптические (коммуникационно-сигнальные). ● Заякоревающие контакты, кроме адгезивного сцепления мембран соседних клеток, со стороны цитоплазмы служат точками опоры для филаментов субмембранного цитоскелета (см. выше). Такие контакты обычно бывают точечными, но иногда образуют сплошную ленту, опоясывая клетку по периметру. В подвижных клетках амёбоидного типа в области т.н. фокальных контактов с помощью особых связующих белков (винкулин и др.) к плазмалемме прицепляются (заякореваются) пучки актиновых микрофиламентов (6-7 нм). С участием миозина они обеспечивают фагоцитоз, сокращение и перемещение клеток по внеклеточному субстрату (матриксу). В эпителиальных клетках такие пучки могут создавать сплошное сократимое кольцо в апикальной части, способствуя образованию эпителиальных складок и трубок. ● Широко распространены контакты заякоревающего типа, называемые десмосомами (клетка – клетка) или полудесмосомами (клетка – матрикс). Они создают прочные межклеточные соединения (белок кадгерин) и, кроме того, фиксируют на себе пучки промежуточных филаментов (10 нм, белок кератин и др.) с помощью белка десмоплакина. Т.о., десмосомы выполняют одновременно контактную и опорную функции. ● Запирающие (плотные) контакты создают на апикальной поверхности эпителиев (сразу под микроворсинками) по всему периметру клеток сетчатую зону слияния мембран, запирая проходы из окружающей среды в межклеточные пространства. ● Специальные интегральные белки от двух мембран контактирующих клеток (окклюдины) плотно склеиваются между собой, не оставляя даже щелей между мембранами. Вокруг каждой клетки создается сплошной пояс непроницаемости. В кишке эпителий ограждается от действия ферментов, в почечных канальцах – от мочи. ● Щелевые контакты и синапсы служат для коммуникации и передачи сигналов между соседними клетками: мышечными, нервными и др. ● Щелевой контакт образуется скоплением множества (от 10 до нескольких тысяч) белковых поровых комплексов – коннексонов, расположенных симметрично в контактирующих мембранах. Коннексон состоит из 6 молекул белка коннектина, которые через очень узкую щель (2-3 нм) фактически склеены с такими же молекулами соседней мембраны. Внутри такого комплекса есть канал сечением 2 нм, по которому проходят малые сигнальные вещества или метаболиты. Так, при развитии организма координируется специализация разных клеток; в сердце синхронизируется сокращение кардиомиоцитов без участия нервных элементов. Синаптические контакты (синапсы) служат для односторонней передачи сигнала между нервными клетками или от нервного окончания на мышцу. Синапсы могут быть химические – с выбросом медиаторов в синаптическую щель, либо электрические – при плотном контакте мембран. ● В растительных тканях целлюлозные клеточные стенки, склеенные межклеточным веществом (срединная пластинка), можно рассматривать как своеобразные механические межклеточные контакты (соединения). При их пропитывании лигнином образуется прочная древесина. ● Между растительными клетками имеются также коммуникационные контакты – плазмодесмы – канальцы, в которых сливаются плазмалеммы и сама цитоплазма соседних клеток, даже цистерны ЭПР. Возможен прямой переток веществ из клетки в клетку. Дополнительные вопросы: Что такое иммунная реакция и толерантность на клеточном уровне? Каково значение этих реакций для поддержания клеточно-тканевого гомеостаза? Показать структурно-функциональное единство элементов поверхностного аппарата и цитоскелета в реакции адгезии фибробласта на субстрате. После дезагрегации и перемешивания в питательной среде клетки почки собираются в агрегаты по тканевому признаку, клетки от оранжевых и серых губок - по видовому (агрегируют клетки одного цвета). Как это объяснить свойствами клеточной поверхности? |