ткани. ТКАНИ. Организация эпителиев Пограничное расположение
 Скачать 1.86 Mb. Скачать 1.86 Mb.
|

|
Организация эпителиев Пограничное расположение Эпителии отделяют организм от внешней среды и от вторичных полостей тела. Эту задачу выполняют пласты эпителия. Образуя непрерывный слой, эпителий отделяет подлежащие ткани от внешней среды и от вторичных полостей тела. Толщина пластов различна. Например, эпидермис кожи имеет толщину до нескольких десятков мкм, тогда как эпителий на поверхности альвеол лёгкого - около 0,2 мкм. Пласт - не единственный тип организации эпителиев. Незначительные межклеточные пространства В эпителии практически нет межклеточного вещества, клетки плотно примыкают одна к другой и связаны с помощью специализированных межклеточных контактов. Эпителиоциты формируют адгезионные (промежуточный, десмосома и полудесмосома), замыкающие (плотный) и коммуникационные (щелевой) контакты. Полярная дифференцировка эпителиальных клеток Базальная и апикальная части клетки отличаются как структурно, так и функционально. Этот признак обязателен для однослойных эпителиев пограничного расположения (на границе внешней и внутренней сред, на поверхности серозных оболочек), а также для эпителиальных клеток, находящихся в тесной связи с кровеносными капиллярами (например, в эндокринных железах, печени). 1)Апикальная часть содержит микроворсинки, стереоцилии, реснички, секреторный материал и участвует в образовании плотных и промежуточных контактов. В эпителиоцитах, транспортирующих глюкозу из апикальной в базальную часть, в плазматическую мембрану апикальной части встроены переносчики глюкозы. В плазматической мембране апикальной части каёмчатых клеток крипт тонкой кишки присутствуют системы транспорта ионов Cl- и Na+ из клетки в просвет органа. В плазмолемме апикальной части клетки преобладают фосфатидилэтаноламин и фосфатидилсерин. 2)Базальная часть содержит различные органеллы. Расположение митохондрий преимущественно в базальной части связано с необходимостью АТФ для встроенных в плазмолемму этой части клетки ионных насосов (например, Na+,K+‑АТФаза). В базальной части клетки присутствуют рецепторы гормонов и факторов роста, транспортные системы ионов и аминокислот. Переносчики глюкозы базальной части (обеспечивающие выход глюкозы из клетки по концентрационному градиенту) отличаются от встроенных в апикальную мембрану. Плазмолемма базальной части содержит преимущественно фосфатидилхолин, сфингомиелин и фосфатидилинозитол. Базальная мембрана Базальная мембрана (базальная пластинка) имеет толщину 20-100 нм, отделяет эпителий от подлежащей соединительной ткани, укрепляет эпителиальный пласт, образуется за счёт эпителия и подлежащей соединительной ткани, содержит коллаген типа IV, ламинин, энтактин и протеогликаны. Эпителиальные клетки прикреплены к базальной мембране с помощью полудесмосом. Через базальную мембрану осуществляется питание эпителия. Отсутствие кровеносных сосудов Питание эпителия, транспорт газов, выведение продуктов метаболизма из эпителия осуществляются путём диффузии веществ через базальную мембрану между эпителием и подлежащими кровеносными сосудами. В эпителиальных злокачественных опухолях (карциномы) целостность базальной мембраны и межклеточных контактов нарушается, и кровеносные сосуды прорастают в эпителиальную опухолевую ткань. Пространственная организация Эпителиальные клетки организованы в ассоциаты на границе внутренней и внешней среды организма, а также во внутренней среде следующим образом: пласт, тяж, островок, фолликул, трубочка, сеть. Пласт. Эпителиальные клетки, формирующие пласты, всегда имеют пограничное положение (например, эпидермис, эпителии слизистой оболочки кожного и кишечного типа, мезотелии). Для клеток однослойного пласта характерна полярная дифференцировка, а многослойные пласты имеют значительные морфологические отличия между эпителиальными клетками разных слоёв. Трубочка - вариант пласта, свёрнутого в трубочку (например, потовые железы, канальцы нефрона. Островок. Эпителиальные островки всегда погружены во внутреннюю среду организма и, как правило, выполняют эндокринную функцию (например, островки поджелудочной железы). Фолликул - имеющий полость островок эпителия. Типичный пример - фолликулы щитовидной железы. Тяж. По принципу анастомозирующих тяжей из эпителиальных гепатоцитов организована паренхима печени. Сеть. В вилочковой железе поддерживающий каркас состоит из отростчатых и контактирующих друг с другом эпителиальных клеток. Способность к регенерации Регенерация выражена у покровных эпителиев и вытекает из их пограничного расположения. Необходимые условия для регенерации - доказанное или предполагаемое наличие стволовых клеток (например, в эпидермисе, эпителии слизистой оболочки трубчатых и полостных органов, мезотелии), возможность репликации ДНК с последующим цитокинезом или без него (например, гепатоциты). У погружённых во внутреннюю среду эпителиальных клеток регенераторные возможности существенно меньше, вплоть до полной невозможности регенерации (например, β-клетки островков поджелудочной железы). Цитокератины Промежуточные филаменты клеток различных эпителиев имеют разные молекулярные формы цитокератина. Более того, в различных анатомических областях одного и того же эпителия могут экспрессироваться различные формы цитокератина. Например, кератиноциты ладони и подошвы синтезируют особые кератины, не встречающиеся в других частях тела. Известно более 20 форм кератина с Mr от 48 до 68 кД; каждая форма кодирована своим геном. По мере дифференцировки эпителиоцитов происходит перепрограммирование синтеза кератинов (например, в эпидермисе). Экспрессия некоторых кератинов - признак появления клеток, достигших состояния терминальной дифференцировки. Так, цитокератин 1 служит маркёром терминальной дифференцировки кератиноцитов. Классификации эпителиальных пластов Однослойные пласты (плоский, кубический, цилиндрический). Все клетки контактируют с базальной мембраной. Однорядный эпителий - ядра клеток расположены в один ряд, т.е. на одинаковом расстоянии от базальной мембраны. Представлен одинаковыми клетками (например, однослойный эпителий канальцев почки). Многорядный - ядра клеток расположены в несколько рядов, т.е. на различном расстоянии от базальной мембраны. Представлен клетками различной величины и формы. Типичный пример однослойного многорядного эпителия - мерцательный эпителий слизистой оболочки воздухоносных путей. Многослойные эпителии подразделяют на многослойный плоский ороговевающий, многослойный плоский неороговевающий и многослойный переходный эпителии. Такие пласты складываются из пролиферативных единиц. Многослойный плоский ороговевающий эпителий (эпидермис) присутствует в коже и имеет роговой слой, состоящий из плотно упакованных роговых чешуек, содержащих ковалентно связанные с плазмолеммой нерастворимые белки. Многослойный плоский неороговевающий эпителий не содержит рогового слоя. Многослойный переходный эпителий. Его поверхностные клетки имеют особую организацию. При растяжении стенки органа поверхностные клетки меняют форму таким образом, что целостность эпителиального пласта не нарушается. Функции эпителиев Транспорт газов (O2 и CO2) через эпителий альвеол лёгких; аминокислот и глюкозы с помощью специальных транспортных белков в эпителии кишки; IgA и других молекул на поверхность эпителиальных пластов. Эндоцитоз, пиноцитоз. Эпителиальные клетки участвуют в пиноцитозе (например, эпителий почечных канальцев) и в опосредуемом рецепторами эндоцитозе (например, поглощение холестерина вместе с ЛНП или трансферрина большинством эпителиальных клеток). Секреция. Экзоцитоз слизи, гормонов, факторов роста, ферментов. Барьерная. Разграничение сред путём образования барьеров из эпителиальных клеток, связанных плотными контактами (например, между эпителиальными клетками слизистой оболочки желудка и кишки). Защита организма от повреждающего действия физических и химических факторов внешней среды. Эпителиальные железы Морфология. Экзокринные железы состоят из секреторных клеток, образующих секреторный (концевой) отдел, и выводного протока. В состав секреторного отдела, кроме железистых (секреторных) клеток, могут входить миоэпителиальные клетки. Они образуют длинные отростки, охватывающие снаружи концевые отделы. Сокращаясь, миоэпителиальные клетки облегчают продвижение секрета в выводной проток. Железистая клетка синтезирует, накапливает, хранит и выделяет секрет. В клетках, вырабатывающих белковый секрет, хорошо развита гранулярная эндоплазматическая сеть, активно функционирует комплекс Гольджи. Гладкая эндоплазматическая сеть выражена в клетках, вырабатывающих небелковые секреты (например, стероидные гормоны). Выводной проток служит для оттока секрета из железы. В крупных железах различают внутридольковые, междольковые, междолевые и главный протоки. Классификация. Железы классифицируют по следующим критериям: форма и ветвление секреторного отдела, ветвление выводного протока, тип секрета В зависимости от формы секреторного отдела различают альвеолярные, трубчатые и смешанные (альвеолярно-трубчатые) железы; в зависимости от ветвления секреторного отдела - разветвлённые и неразветвлённые. Форма выводного протока определяет деление желёз на простые (проток не ветвится) и сложные (проток ветвится). От типа секрета зависит разделение на серозные (белковые), слизистые и белково-слизистые железы. Способ секреции. Различают несколько вариантов отделения секрета Эккриновый (мерокриновый) - выделение секрета путём экзоцитоза (слюнные железы). Апокриновый - отделение секрета вместе с фрагментом апикальной части секреторной клетки (молочнаяжелеза).Голокриновый - полное разрушение секреторной части(сальная) 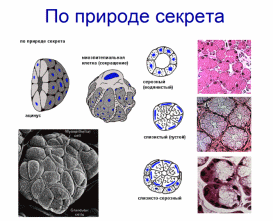 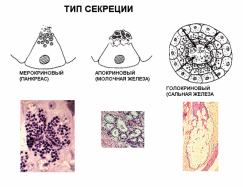 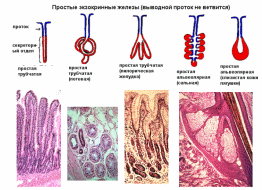 Рис. 30. Схема межклеточных соединений в эпителиях: А - область расположения комплекса межклеточных соединений (выделена рамкой): 1 - эпителиоцит: 1.1 - апикальная поверхность, 1.2 - латеральная поверхность, 1.2.1 - комплекс межклеточных соединений, 1.2.2 - пальцевидные соединения (интердигитации), 1.3 - базальная поверхность; 2 - базальная мембрана. Б - вид межклеточных соединений на ультратонких срезах (реконструкция): 1 - плотное (замыкающее) соединение; 2 - опоясывающая десмосома (адгезивный поясок); 3 - десмосома; 4 - щелевое соединение (нексус). В - трехмерная схема строения межклеточных соединений: 1 - плотное соединение: 1.1 - внутримембранные частицы; 2 - опоясывающая десмосома (адгезивный поясок): 2.1 - микрофиламенты, 2.2 - межклеточные адгезивные белки; 3 - десмосома: 3.1 - десмосомная пластинка (внутриклеточное десмосомное уплотнение), 3.2 - тонофиламенты, 3.3 - межклеточные адгезивные белки; 4 - щелевое соединение (нексус): 4.1 - коннексоны 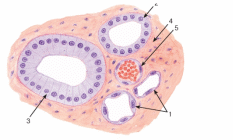 Рис. 33. Однослойные плоский, кубический и столбчатый (призматический) эпителии (мозговое вещество почки) Окраска: гематоксилин-эозин 1 - однослойный плоский эпителий; 2 - однослойный кубический эпителий; 3 - однослойный столбчатый эпителий; 4 - соединительная ткань; 5 - кровеносный сосуд 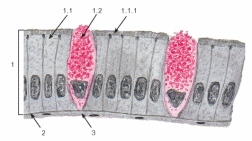 Рис. 34. Однослойный столбчатый каемчатый (микроворсинчатый) эпителий (тонкая кишка) Окраска: железный гематоксилин-муцикармин 1 - эпителий: 1.1 - столбчатый каемчатый (микроворсинчатый) эпителиоцит (энтероцит), 1.1.1 - исчерченная (микроворсинчатая) каемка, 1.2 - бокаловидный экзокриноцит; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань 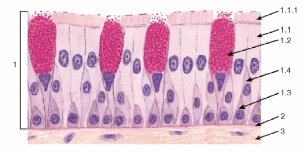 Рис. 36. Однослойный многорядный столбчатый реснитчатый (мерцательный) эпителий (трахея) 1 - эпителий: 1.1 - реснитчатый эпителиоцит, 1.1.1 - реснички, 1.2 - бокаловидный экзокриноцит, 1.3 - базальный эпителиоцит, 1.4 - вставочный эпителиоцит; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань 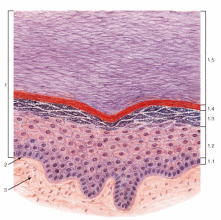 Рис. 38. Многослойный плоский ороговевающий эпителий (эпидермис толстой кожи) Окраска: гематоксилин-эозин 1 - эпителий: 1.1 - базальный слой, 1.2 - шиповатый слой, 1.3 - зернистый слой, 1.4 - блестящий слой, 1.5 - роговой слой; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань 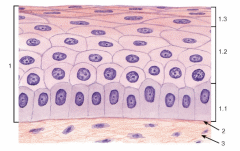 Рис. 39. Многослойный плоский неороговевающий эпителий (роговица) 1 - эпителий: 1.1 - базальный слой, 1.2 - промежуточный слой, 1.3 - поверхностный слой; 2 - базальная мембрана; 3 - рыхлая волокнистая соединительная ткань ТКАНИ ВНУТРЕННЕЙ СРЕДЫ 1)КРОВЬ Жидкое межклеточное вещество (плазма) и взвешенные в нём клетки - два основных компонента крови. Свернувшаяся кровь состоит из тромба (сгустка), включающего форменные элементы и некоторые белки плазмы, и сыворотки - прозрачной жидкости, сходной с плазмой, но лишённой фибриногена. ПЛАЗМА: состоит из воды (90%), органических (9%) и неорганических (1%) веществ. Белки составляют 6% всех веществ плазмы Белки плазмы: 1)Белки системы свёртывания крови. Различают коагулянты и антикоагулянты. Обе группы белков обеспечивают равновесие между процессами формирования и разрушения тромба. Коагулянты участвуют в формировании тромба. К коагулянтам, например, относят фибриноген (синтезируется в печени и при гемокоагуляции превращается в фибрин). Антикоагулянты - компоненты фибринолитической системы, препятствующие свёртыванию (например, антитромбин III). В плазме крови присутствуют прокоагулянтные микрочастицы, содержащие тканевый фактор. Микрочастицы могут происходить из тромбоцитов, лейкоцитов, эндотелиальных клеток, гладкомышечных клеток и содержат тканевый фактор, необходимый для тромбообразования. 2)Белки, участвующие в воспалительных и иммунных реакциях. К этой группе относят Ig, белки комплемента, α2-макроглобулины. Белки комплемента (C1-C9) участвуют в неспецифической защите (врождённый иммунитет) и инициируют реакции воспаления. α2-Макроглобулины плазмы - гликопротеины, сходные с белками комплемента по функциям и структуре. Эти антипротеазы нейтрализуют токсическое действие ферментов, выделяющихся при реакциях воспаления и повреждения, в частности, ингибируют протеазы, связывают цитокины (ИЛ1, ИЛ6) и факторы роста. 3)Транспортные белки - альбумины, аполипопротеины, трансферрин, гаптоглобин, церулоплазмин, транскортин, транскобаламины и др. Альбумины (Mr ≈ 60-65 кД) составляют основную часть белков плазмы, связывают и переносят мало- или нерастворимые метаболиты (например, жирные кислоты), многие гормоны, переносят билирубин в печень для последующей экскреции. КЛЕТКИ КРОВИ Эритроциты Продолжительность жизни (время циркуляции в крови) - 100-120 сут. Разброс размеров эритроцитов - анизоцитоз, клетки диаметром >9 мкм - макроциты, <6 мкм - микроциты. При ряде заболеваний крови изменяются и размеры (анизоцитоз) и форма эритроцитов (пойкилоцитоз). Ретикулоциты - незрелые эритроциты, поступающие в кровоток из костного мозга. Они содержат рибосомы, митохондрии, комплекс Гольджи. Окончательная дифференцировка в эритроциты происходит в течение 24-48 ч после выхода в кровоток. Количество поступающих в кровоток ретикулоцитов в норме равно количеству удаляемых эритроцитов. Ретикулоциты составляют около 1% всех циркулирующих красных клеток крови. Плазмолемма и примембранный цитоскелет Клеточная мембрана эритроцита довольно пластична, что позволяет клетке деформироваться и легко проходить по узким капиллярам диаметром 3-4 мкм. Главные трансмембранные белки эритроцита - белок полосы 3, гликофорины, транспортёр глюкозы GLUT1. Белок полосы 3 (рис. 6-1) совместно с белками примембранного цитоскелета (спектрин, анкирин, актин, белок полосы 4.1) обеспечивает поддержание формы эритроцита в виде двояковогнутого диска. Белок полосы 3 - полифункциональный мембранный гликопротеин, участвует в переносе анионов Cl- и НСО3- через плазмолемму (анионный транспортёр), имеет участки связывания с анкирином, удерживающим спектрин-актиновую сеть непосредственно под мембраной эритроцита. Анкирин- посредник между белком полосы 3 и нитями спектрина. Спектрин. Две изоформы белка (α и β) образуют димеры, которые, в свою очередь, соединяются "конец в конец", формируя тетрамеры - гибкие нити примембранного цитоскелета. Концы нитей спектрина связываются с глобулярным белком полосы 4.1. Белок полосы 4.1 взаимодействует со спектриновыми и актиновыми нитями. Актин. В составе примембранного цитоскелета каждая актиновая нить прикрепляется к цепочке из нескольких молекул белка полосы 4.1. Таким образом, актиновые нити вместе с гетеродимерами спектрина и глобулярным белком полосы 4.1 образуют примембранный скелет, который через анкирин связан с интегральным белком полосы 3. Гликофорины А, B, C, D, Е - трансмембранные сиалогликопротеины, их полисахаридные цепи содержат антигенные детерминанты, позволяющие типировать группы крови по системе MNS (43 известных фенотипа). Транспортёр глюкозы GLUT1 обеспечивает инсулинзависимый трансмембранный перенос глюкозы в цитоплазму эритроцита (облегчённая диффузия). Глюкоза является основным источником энергии в зрелых эритроцитах. Путём гликолиза и/или гексозомонофосфатного шунта глюкоза расщепляется с образованием АТФ. Гемоглобин Эмбриональные Hb появляются у 19-дневного эмбриона, присутствуют в эритроидных клетках в первые 3-6 мес беременности. Фетальный Hb (HbF) появляется на 8-36 нед беременности и составляет 90-95% всего Hb плода. Дефинитивные Hb составляют основное количество Hb эритроцитов взрослого человека Гем построен из четырёх молекул пиррола, образующих порфириновое кольцо, в центре которого находится атом железа. Гем связывает кислород и высвобождает электроны для ферментов, катализирующих окислительно-восстановительные реакции. Недостаточность ферментов, необходимых для синтеза гема, - причина снижения количества Hb и развития гипохромной анемии. Оксигемоглобин. В лёгких при повышенном pO2 Hb присоединяет O2, образуя оксигемоглобин. При включении в гем трёхвалентного железа образуется метгемоглобин. Метгемоглобин - Hb с Fe3+ прочно связывает O2, так что отделение последнего затруднено. Это приводит к нарушениям газообмена в тканях (метгемоглобинемия). Образование метгемоглобина в эритроцитах может быть наследственным или приобретённым. В последнем случае это результат воздействия на эритроциты сильных окислителей. К ним относят нитраты и неорганические нитриты, лекарственные средства (сульфаниламиды) и местные анестетики (лидокаин). Карбоксигемоглобин. Hb легче, чем с O2, связывается с окисью углерода СО (угарный газ), образуя карбоксигемоглобин. Гликозилированный гемоглобин (AIa1, AIa2, AIb, AIc). В норме Hb, связавший d-глюкозу, составляет менее 4% общего Hb крови. Содержание гликозилированного Hb увеличено при сахарном диабете пропорционально содержанию глюкозы в плазме крови. Ферменты-гликолиз (расщепление глюкозы с образованием АТФ); газообмен; сохранение структуры плазмолеммы и формы клетки; транспорт электролитов через цитолемму и обеспечение их оптимальной внутриклеточной концентрации; поддержание в восстановленной форме Fe2+ в молекуле Hb; сохранение Hb в растворённом состоянии. Газообмен: Транспорт кислорода. Одна молекула Hb связывает четыре молекулы O2 с образованием оксигемоглобина. Транспорт углекислого газа. О2 транспортируется кровью преимущественно в виде бикарбонат иона (НСО3-). В тканях СО2 путём пассивного транспорта диффундирует в кровеносные капилляры. В эритроцитах под влиянием карбоангидразы CO2 взаимодействует с H2O с образованием угольной кислоты (H2CO3), диссоциирующей на HCO3- и H+ (Н2О ± СО2 ↔ H+ ± НСО3-). Через анионный транспортёр (белок полосы 3) бикарбонат ион (НСО3-) в обмен на анион Cl- выходит в плазму, а Н+ связывается с дезоксигемоглобином. В капиллярах лёгких НСО3- транспортируется из плазмы обратно в эритроциты, где карбоангидраза катализирует реакцию Н+ ± НСО3- с образованием СО2 и Н2О. Гибель эритроцитов в селезёнке, а также в печени и костном мозге. снижается обмен веществ, нарушается форма, происходит деградация белков, появляются новые Аг. Такие стареющие клетки распознаются макрофагами и фагоцитируются. В эритроцитах так называемый "Аг старения" представлен в виде деградированного белка полосы 3. При разрушении Hb глобины расщепляются до аминокислот, а из гема освобождаются ионы железа, СО и вердоглобин, который переходит в биливердин, восстанавливающийся в билирубин. Свободный билирубин в комплексе с альбумином транспортируется в печень. В гепатоцитах происходит конъюгация билирубина с глюкуроновой кислотой; в составе жёлчи связанный билирубин поступает в кишечник. Обмен железа Основное количество железа в организме входит в состав гема (Hb, миоглобин, цитохромы). Некоторое количество железа находится в лабильном состоянии в плазме в связи с трансферрином. Большая часть железа, необходимого для синтеза гема, извлекается из разрушенных эритроцитов. При разрушении эритроцитов в селезёнке, печени и костном мозге высвобождаемое из гема железо транспортируется трансферрином в костный мозг, а часть железа включается в состав ферритина (в гепатоцитах) и гемосидерина (в макрофагах печени, костного мозга и селезёнки). Только 5% железа для эритропоэза поставляется извне с пищей. .Лейкоциты В цитоплазме лейкоцитов находятся гранулы - специфические (вторичные) и азурофильные (первичные, сходные с лизосомами). Гранулоциты (нейтрофилы, эозинофилы, базофилы) содержат специфические и азурофильные гранулы. В цитоплазме агранулоцитов (моноцитов, лимфоцитов) присутствуют только азурофильные гранулы. Для гранулоцитов характерно дольчатое сегментированное ядро разнообразной формы, в связи с чем их называют полиморфноядерными лейкоцитами. Лимфоциты и моноциты имеют несегментированное ядро, их называют мононуклеарными лейкоцитами. Лейкоциты участвуют в защитных реакциях: уничтожают микроорганизмы, захватывают инородные частицы и продукты распада тканей, осуществляют реакции гуморального и клеточного иммунитета. Лейкоциты используют кровоток как средство пассивного транспорта. Они содержат сократительные белки (актин, миозин), что позволяет им выходить из кровеносных сосудов Лейкоциты выходят в окружающую сосуд соединительную ткань, проникая между эндотелиальными клетками (диапедез) и разрушая секретируемыми ими ферментами базальную мембрану эндотелия. Последующую направленную миграцию лейкоцитов (хемокинез, хемотаксис) контролируют различные вещества (в т.ч. хемоаттрактанты). НЕЙТРОФИЛЫ: образуются в костном мозге, выходят в кровоток и циркулируют 8-12 ч. Продолжительность жизни - около 8 сут Выделяют три пула нейтрофилов: циркулирующий, пограничный и резервный. палочкоядерные нейтрофилы - незрелые формы клеток с подковообразным ядром. Строение Ядро зрелой клетки - сегментоядерного нейтрофила - состоит из 3-5 сегментов, соединённых тонкими перемычками, хроматин сильно конденсирован. В нейтрофилах у женщин один из сегментов ядра содержит вырост в форме барабанной палочки - тельце Барра. Тельце Барра заметно у 3% нейтрофилов в мазке крови женщин Много гликогена.(энергия путем гликолиза) Мало митохондрий и органелл для синтеза Б В плазмолемме рецепторы молекул адгезии, цитокинов, колониестимулирующих факторов, опсонинов Азурофильные гранулы (красновато-пурпурного цвета на окрашенном мазке) - специализированные лизосомы нейтрофилов - являются основным депо лизосомных кислотозависимых гидролаз,содержат различные белки, разрушающие компоненты внеклеточного матрикса и обладающие антибактериальной активностью. В гранулах присутствуют миелопероксидаза, протеиназа 3 (миелобластин), азуроцидин, эластаза, катепсины A, C, D, E, G, L, дефензины, катионные белки, лизоцим, арилсульфатаза. Миелопероксидаза (MPO) составляет 2-4% массы нейтрофила, катализирует образование хлорноватистой кислоты HOCl и других токсических агентов, значительно усиливающих бактерицидную активность нейтрофилов. Протеиназа 3 расщепляет эластин. Азуроцидин- сериновая протеаза с антибактериальной активностью. Эластаза нейтрофилов -протеолиз коллагена типа IV, эластина и других белков внеклеточного матрикса при дегенеративных и воспалительных заболеваниях, разрушает наружный мембранный белок A грамотрицательных бактерий, модифицирует функции моноцитов, гранулоцитов, NK-клеток. Дефензины повышают проницаемость мембран грамотрицательных бактерий. Катепсин C-отщеплении дипептидов Катепсин G при нейтральном рН убивает грамположительные и грамотрицательные бактерии Катепсин L большую роль во внутриклеточном катаболизме белка. вовлечён в различные патологические процессы Лизоцим разрушает пептидогликан стенки бактерии, что приводит к лизису клетки. Повышающий проницаемость бактерий белок BPI Специфические гранулы содержат белки, обладающие бактериостатическими свойствами и разрушающие внеклеточный матрикс: лактоферрин, витамин B12‑связывающие белки, лизоцим, металлопротеиназы (желатиназу и коллагеназу, липокалин), щелочную фосфатазу, катионные белки (кателицидин). Лактоферрин бактериостатические свойства; связывает также свободные радикалы, продуцируемые нейтрофилами и повреждающие как сами клетки, так и окружающие ткани. Металлопротеиназы. Кателицидин -связывается с бактериальными липополисахаридами Кателицидин не только проявляет мощную бактерицидную активность, является хемоаттрактантом для клеток иммунной защиты. Рецепторы. молекул адгезии, цитокинов, колониестимулирующих факторов опсонинов, медиаторов воспаления и бактериальных продуктов. Активация и хемотаксис. Биологически активные соединения различного происхождения воздействуя на нейтрофилы,стимулируют их активность. +являются хемоаттрактантами, по градиенту концентрации которых происходит миграция нейтрофилов. Адгезия. Активированный нейтрофил прикрепляется к эндотелию сосуда в участке, ближайшем к месту воспаления. Адгезия нейтрофилов к эндотелию опосредована селектинами молекулами адгезии. Плотная адгезия опосредуется молекулами адгезии ICAM эндотелия, связывающимися с интегринами нейтрофилов. После связывания интегринов с ICAM нейтрофил останавливается, меняет форму (распластывается) и проходит между эндотелиальными клетками, взаимодействуя с ними через молекулу адгезии PECAM-1Для разрушения базальной мембраны нейтрофил секретирует гидролазы. Миграция. После прикрепления к эндотелию и выхода из сосуда нейтрофилы увеличиваются в размерах, образуя широкий головной конец (ламеллоподия) и суженную заднюю часть (уроподия). Нейтрофил, продвигая вперёд ламеллоподию, мигрирует к источнику хемоаттрактанта. При этом гранулы перемещаются к головному концу, происходит экзоцитоз содержимого гранул. В изменении формы активированного нейтрофила участвует кальпаин -нелизосомная цистеиновая Ca2+-связывающая протеаза, присутствующая в цитоплазме клетки. Высокая активность кальпаина в покоящихся нейтрофилах угнетает образование псевдоподий и миграцию клеток. Подавление активности (ингибирование) кальпаина способствует поляризации и миграции нейтрофила. Кальпаин регулирует направленную миграцию нейтрофила по градиенту концентрации хемоаттрактанта, модулируя активность Rho ГТФаз Cdc42 и Rac. Функция фагоцитоз опсонизированных микроорганизмов. Фагоцитоз и последующее переваривание материала происходят параллельно с образованием метаболитов арахидоновой кислоты и респираторным взрывом. Распознавание Специфическое распознавание подлежащего фагоцитозу объекта осуществляется с помощью рецепторов к опсонинам Опсонизация резко усиливает фагоцитарную активность нейтрофилов. Связывание рецепторов опсонинов нейтрофила вызывает в клетке респираторный взрыв, который, в свою очередь, запускает процесс фагоцитоза. Неопсонизированные микроорганизмы устойчивы к фагоцитозу и поэтому весьма патогенны. Нейтрофил способен распознавать бактерии без предварительной их опсонизации через Toll-подобные рецепторы TLR Респираторный взрыв Нейтрофилы в течение первых секунд после стимуляции резко увеличивают поглощение кислорода и быстро расходуют значительное его количество. Это явление известно как респираторный (кислородный) взрыв. При этом образуются токсичные для микроорганизмов соединения кислорода. образуется H2O2 и свободный радикал OH-. Фагоцитоз Фагоцитоз Распознавание-инвагинация мембраны-фагосома-фагосома+лизосома=фаголизосома-уничтожение бактерии и разрушение захваченного материала(. Для этого в фаголизосому поступают: лизоцим, катепсин, эластаза, лактоферрин, дефензины, катионные белки; миелопероксидаза; супероксид О2- и гидроксильный радикал ОН-, образующиеся (наряду с Н2О2) при респираторном взрыве.)- нейтрофил погибает-гнойные клетки(+ погибшие макрофаги,бактерии,тк.ж-ть) Эозинофилы мигрируют в ткани, контактирующие с внешней средой (слизистые оболочки дыхательных и мочеполовых путей, кишечника) 1-5% лейкоцитов, циркулирующих в крови Продолжительность жизни - предположительно 8-14 суток. СТРОЕНИЕ Ядро из двух крупных сегментов, соединённых тонкой перемычкой (сегментоядерный эозинофил). Цитоплазма содержит хорошо развитую гранулярную эндоплазматическую сеть, небольшое количество цистерн гладкой эндоплазматической сети, скопления рибосом, отдельные митохондрии и много гликогена. Специфические гранулы. В цитоплазме с выраженной ацидофилией (красно-оранжевые) Рецепторы.-мембранные Метаболиты арахидоновой кислоты, включая лейкотриен LTС4 и PAF. Активация. Факторы-ИЛ2, ИЛ3, ИЛ5; колониестимулирующие факторы GM-CSF, G-CSF; факторы, происходящие из эндотелия; PAF; TNF; ИФН; белки комплемента, паразитарные факторы и др. Хемотаксис. Активированные эозинофилы перемещаются по градиенту многих факторов хемотаксиса - бактериальных и паразитарных факторов, фактора хемотаксиса эозинофилов (ECF) и гистамина из тучных клеток, белков комплемента, эотаксина, ИЛ5. ФУНКЦИИ Уничтожение паразитов. Активированный эозинофил выделяет содержимое гранул и липидные медиаторы- повреждающее действие на паразитов. Особенно эффективен главный щелочной белок MBP специфических гранул. Участие в аллергических реакциях. Содержимое гранул блокирует дегрануляцию тучных клеток, инактивирует гистамин и лейкотриен LTС4. +выделяют ингибитор, блокирующий дегрануляцию тучных клеток. Медленно реагирующий фактор анафилаксии (SRS-A), секретируемый базофилами и тучными клетками, также ингибируется активированными эозинофилами. Базофилы В крови базофилы находятся 1-2 сут. Размер - 10-12 мкм. Уплотнённое ядро состоит из нечётко выраженных трёх долек, изогнуто в виде буквы S. В цитоплазме имеются все виды органелл, свободные рибосомы и гликоген. Специфические гранулы довольно крупные (0,5-1,2 мкм), окрашиваются метахроматически (от красновато-фиолетовых до интенсивно-фиолетовых). Имеют разнообразную, чаще овальную или округлую форму с плотным содержимым. В гранулах содержатся различные ферменты и медиаторы. К наиболее значимым из них можно отнести гепаринсульфат (гепарин), гистамин, серотонин, нейтральные протеазы триптазу и химазу, медиаторы воспаления (например, SRS-A, ECF), лизосомные гидролазы. Функция. МИГРИРУЮТ в очаги воспаления и участвуют в аллергических реакциях. Активация и дегрануляция базофилов происходит при попадании в организм аллергена и опосредована IgE. Базофилы имеют высокоаффинные поверхностные рецепторы к Fc-фрагментам IgE. IgE синтезируют плазматические клетки при попадании в организм Аг. Параллельно образуются метаболиты арахидоновой кислоты. Моноциты 2-9% всех лейкоцитов циркулирующей крови. Крупное, эксцентрично расположенное подковообразное ядро имеет пятнистый вид из-за неравномерно конденсированного хроматина. Бледная голубовато-серая (на окрашенном мазке) цитоплазма включает многочисленные лизосомы, содержащие кислые гидролазы, арилсульфатазу, катепсин C, кислую фосфатазу, пероксидазу, разные вакуоли, большое количество рибосом и полирибосом, комплекс Гольджи, мелкие удлинённые митохондрии. Образуются в костном мозге, выходят в кровоток и циркулируют около 2-4 сут. Рецепторы. В мембрану моноцита встроены рецепторы Fc-фрагмента Ig, белков комплемента, цитокинов, медиаторов воспаления, бактериальных продуктов, холинорецепторы, адренорецепторы. Активациямоноцитов. Различные вещества, образующиеся в очагах воспаления и разрушения ткани, - агенты хемотаксиса и активации моноцитов. В результате активации увеличивается размер клетки, усиливается обмен веществ, моноциты выделяют биологически активные вещества (ИЛ1, ИЛ6, TNF-α, M‑CSF, GM‑CSF, sCD14, простагландины, ИФН, факторы хемотаксиса нейтрофилов). Функция. фагоцитоз. Моноциты фагоцитируют опсонизированные частицы. В их переваривании участвуют лизосомные ферменты моноцитов, а также формируемые внутриклеточно H2O2, OH-, O2-. Активированные моноциты/макрофаги продуцируют эндогенные пирогены. Эндогенные пирогены - полипептиды, запускающие метаболические изменения в центре терморегуляции (гипоталамус), что приводит к повышению температуры тела. К эндогенным пирогенам относят ИЛ1, ИЛ6, ИЛ8, TNF-α, α-ИФН. Образование эндогенных пирогенов моноцитами/макрофагами (а также рядом других клеток) вызывают экзогенные пирогены (белки микроорганизмов, бактериальные токсины). Лимфоциты Выделяют следующие типы: В-лимфоциты, Т-лимфоциты и NK-клетки. Т- и В‑лимфоциты морфологически относят к малым (большинство лимфоцитов в кровотоке). Они имеют округлое с небольшими выемками ядро, содержащее конденсированный хроматин, цитоплазма образует узкое кольцо вокруг ядра. Большие, а также средние лимфоциты крови - активированные Аг B-лимфоциты, дифференцирующиеся в плазматические клетки; к большим лимфоцитам также относят NK-клетки. Ядро крупное и относительно плотное. В цитоплазме немного лизосом, небольшое количество митохондрий, рудиментарный комплекс Гольджи, минимум эндоплазматической сети и сравнительно большое количество свободных рибосом. Клетка образует короткие отростки (микроворсинки), более многочисленные у B-лимфоцитов. B-лимфоциты составляют менее 10% лимфоцитов крови. вырабатывают против конкретных Аг соответствующие АТ. T-лимфоциты (80% и более). реагируют на конкретные ,участие в клеточном и гуморальном иммунитете: уничтожают аномальные клетки своего организма, участвуют в аллергических реакциях, отторжении чужеродного трансплантата. CD4+-лимфоциты. (Т-хелперы) поддерживают пролиферацию и дифференцировку В-лимфоцитов, стимулируют образование цитотоксических Т-лимфоцитов. CD4+-лимфоциты подразделяют на две разновидности: Th1 и Th2. Лимфоциты обеих разновидностей поддерживают пролиферацию В-лимфоцитов и секрецию ими Ig, а также секретируют ИЛ3, GM-CSF и TNF-α. CD8+-лимфоциты [цитотоксические Т-лимфоциты (TC)]. В ходе иммунного ответа эти клетки распознают Аг вместе с молекулой MHC I, непосредственно участвуют в реакциях клеточной цитотоксичности. Рецептор Т-лимфоцитов. Каждый клон Т-лимфоцитов производит и содержит рецептор строго одной специфичности, т.е. взаимодействующий только с одним Аг. Функция. участие в клеточном иммунитете. Т-лимфоциты уничтожают собственные клетки, участвуют в реакциях гиперчувствительности замедленного типа, отторгают чужеродный трансплантат. Реакции клеточной цитотоксичности (клеточно-опосредованный цитолиз). В этих реакциях участвуют CD8+-лимфоциты. узнают, связывают и лизируют клетки-мишени. Специальные цитолитические гранулы содержат перфорин, с помощью которого киллеры убивают клетки-мишени. Перфорин- главный цитолитический белок (Mr 70-75 кД) гранул цитотоксических Т-лимфоцитов и NK-клеток; образует трансмембранные каналы в клетках-мишенях. NK-клетки - лимфоциты, лишённые характерных для Т- и В‑клеток поверхностных детерминант. около 5-10% всех циркулирующих лимфоцитов, содержат цитолитические гранулы с перфорином, уничтожают трансформированные, инфицированные вирусами и чужеродные клетки. Идентификация клеток-мишеней не связана с необходимостью узнавания NK-клетками белков MHC на поверхности клеток-мишеней, как это происходит в случае цитотоксических T-лимфоцитов (T-киллеров). При активации (например, под влиянием ИЛ2) NK-клетки приобретают способность к пролиферации. Тромбоциты фрагменты цитоплазмы находящихся в красном костном мозге мегакариоцитов. Количество тромбоцитов в циркулирующей крови - 190-405×109/л, размер - 3-5 мкм. Две трети кровяных пластинок циркулирует в крови, остальные депонируются в селезёнке. Продолжительность жизни - 8 суток. Старые и дефектные тромбоциты фагоцитируются в селезёнке, печени и костном мозге. Строение Плазматическая мембрана. окружён толстым слоем гликокаликса, богатым кислыми гликозаминогликанами. Гликокаликс образует фибриллярные мостики между мембранами соседних тромбоцитов при их агрегации. В составе гликокаликса присутствуют Ca2+ и АДФ, усиливающие адгезию и агрегацию при образовании тромба. Плазматическая мембрана содержит гликопротеины, выполняющие роль рецепторов адгезии и агрегации Цитоплазма на окрашенном мазке - пурпурная и зернистая. Тромбоциты содержат в большом количестве митохондрии, элементы комплекса Гольджи и рибосомы, а также гранулы гликогена и ферменты для аэробного и анаэробного дыхания. Периферическая часть цитоплазмы содержит актин, миозин, гельзолин и другие контрактильные белки, участвующие в округлении тромбоцита и ретракции тромба. Имеются также пучки микротрубочек, циркулярно расположенные под плазмолеммой. Эти микротрубочки необходимы для сохранения овальной формы тромбоцита. В цитоплазме рассеяны узкие, неправильной формы мембранные трубочки, составляющие плотную тубулярную системуПо периферии тромбоцита расположены анастомозирующие краевые мембранные канальцы, открывающиеся во внеклеточную среду; их мембраны связаны с элементами цитоскелета. Система этих канальцев участвует в секреции содержимого α-гранул. Кроме α-гранул, тромбоциты содержат ещё 3 типа гранул - δ-, λ-гранулы и микропероксисомы. |