ОРГАНИЗАЦИЯ ЛАБОРАТОРНОЙ. Организация лабораторной
 Скачать 399.79 Kb. Скачать 399.79 Kb.
|

|
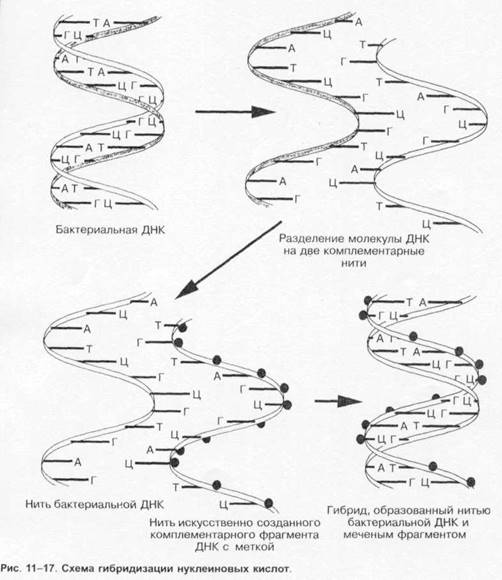
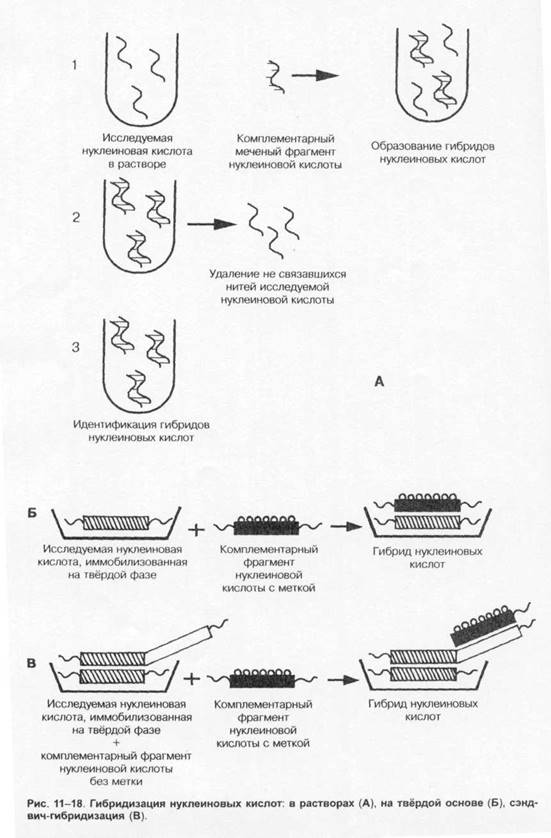 |
Серологические методы
Классические серологические реакции применяют для выявления бактериальных AT, а также для выявления Аг, особенно для идентификации бактериальных Аг. Среди современных методов наибольшее распространение нашли методы твердофазного ИФА и латекс-агглютинации.
Аллергологические методы
Сенсибилизирующей активностью обладает ограниченное количество бактериальных Аг. Поэтому метод кожных проб применяют лишь при диагностике туберкулёза, сапа, мелиоидоза, бруцеллёза и туляремии.
Биологические методы
Выделение патогенных бактерий от заражённых животных имеет большую диагностическую ценность, особенно при контрольном применении иммунных сывороток. Цель подобных манипуляций — уменьшение времени проведения бактериологических исследований.
• При диагностике инфекций, вызванных эффектами токсина (например, ботулизма или сибирской язвы), материал, предположительно содержащий возбудитель и токсин, помещают в физиологический раствор, а затем фильтруют через бумажные фильтры, натёртые тальком (последний хорошо адсорбирует токсин). Смывами с фильтров заражают чувствительных животных.
• При диагностике инфекций, обусловленных различными патогенными свойствами самого возбудителя, лабораторных животных заражают микробной взвесью.
• Для диагностики бактериальных инфекций используют различных животных, так как проявляют видовую восприимчивость к различным этиологическим агентам.
Мыши чувствительны к пневмококкам, нейссериям, пастереллам, клостридиям, листериям, возудителям сибирской язвы, туляремии, чумы, ботулизма, столбняка, коклюша и мелиоидоза
Крысы чувствительны к возбудителям туберкулёза (бычьего типа), мелиоидоза и др.
Морские свинки чувствительны к возбудителям туберкулёза (человеческого типа), дифтерии, сапа, чумы, бруцеллёза, туляремии, холеры, газовой гангрены, ботулизма, псевдотуберкулёза и др.
Кролики чувствительны к стафилококкам, стрептококкам, нейссериям, Mycobacterium bovis , возбудителям газовой гангрены, сибирской язвы, ботулизма, столбняка и др.
Кошки. Животных заражают стафилококками, возбудителями сапа, коклюша и др.
Обезьяны. Их заражают шигеллами, листериями, сальмонеллами, возбудителями мелиоидоза,коклюша и др.
Птицы. Кур и голубей используют для диагностики туберкулёза (птичьего типа), пастереллёза, риносклеромы и др.
МЕТОДЫ ОБНАРУЖЕНИЯ ВИРУСОВ
Лабораторные методы при диагностике вирусных инфекций включают:
• выделение и идентификацию возбудителя;
• обнаружение и определение титров противовирусных AT;
• обнаружение Аг вирусов в образцах исследуемого материала;
• микроскопическое исследование препаратов исследуемого материала.
Забор материала. При заборе материала для исследований необходимо выполнять следующие условия:
• образцы следует отбирать как можно раньше либо с учётом ритма циркуляции возбудителя;
• материал следует отбирать в объёме, достаточном для всего комплекса исследований;
• образцы следует доставлять в лабораторию незамедлительно (!), при относительно кратковременной транспортировке (не более 5 сут) образцы сохраняют на льду, при более длительной — при температуре -50 °С.
Выделение и культивирование вирусов
Выделение и идентификация возбудителя — «золотой стандарт» в диагностике вирусных инфекций.
Культуры клеток
Вирусы размножаются только в живых клетках, и выделение возбудителя в заражённой культуре клеток — один из основных методов диагностики вирусных инфекций. Поскольку большинство патогенных вирусов отличает тканевая и типовая специфичность, то почти к каждому вирусу можно подобрать соответствующие клеточные или тканевые культуры, а также создать стандартные условия культивирования (наличие клеток одного типа). Размножение вируса обеспечивают чувствительные (пермиссивные) клетки. Поэтому при выделении неизвестного возбудителя проводят одномоментное заражение 3-4 культур клеток, предполагая, что одна из них может оказаться пермиссивной. Культуры клеток получают диспергированием соответствующих органов и тканей, но чаще используют эмбриональные ткани (человека и животных) либо трансформированные опухолевые клетки. При помещении на соответствующую плоскую поверхность клеточные культуры обычно растут в виде монослоя.
Первично-трипсинизированные культуры. Суспензии клеток получают гомогенизированием соответствующих тканей, предварительно обработанных трипсином. Культуры часто представлены клетками смешанного типа и не подлежат повторному культивированию. Жизнеспособность таких культур составляет 2-3 нед.
Полуперевиваемые линии клеток представлены диплоидными клетками человека и животных. Культуры ограниченно пригодны к повторному диспергированию и росту (как правило, не более 20-30 пересевов), сохраняя при этом жизнеспособность и не подвергаясь спонтанной трансформации.
Перевиваемые линии клеток (гетероплоидные культуры) представлены клетками, подвергнутыми длительному культивированию и спонтанным трансформациям. Культуры способны к многократному диспергированию и перевиванию. Работа с ними менее трудоёмка по сравнению с приготовлениями первичных культур; перевиваемые клетки относительно одинаковы по своей морфологии и стабильны по свойствам.
Культуры органов
Не все виды клеток способны расти в виде монослоя, в некоторых случаях поддержание дифференцированных клеток возможно только в культуре органа. Обычно это суспензия ткани, обладающей специализированной функцией, также обозначаемая как культура переживающей ткани.
Куриные эмбрионы
Куриные эмбрионы (рис. 1-20) — практически идеальные модели для культивирования некоторых вирусов (например, гриппа и кори). Замкнутая полость эмбриона препятствует проникновению микроорганизмов извне, а также развитию спонтанных вирусных инфекций. Эмбрионы применяют для первичного выделения вирусов из патологического материала; для пассирования и сохранения их, а также для получения необходимых количеств вируса. Некоторые возбудители (например, герпесвирусы) вызывают характерные изменения (по ним можно распознавать заболевание). Заражение проводят на хорион-аллантоисную оболочку, в амниотическую или аллантоисную полость либо в желточный мешок.
Заражение на хорион-аллантоисную мембрану. Обычно используют 10-12-суточные эмбрионы. Яйца просматривают в проходящем свете, отмечают локализацию воздушного мешка и выбирают область без сосудов. Осторожно удаляют фрагмент скорлупы, освобождают наружную оболочку и отслаивают её осторожным надавливанием. Затем делают отверстие у края воздушного мешка. При отсосе через это отверстие хорион-аллантоисная оболочка отслаивается от наружной оболочки. На неё наносят исследуемый материал, свободный от бактерий и простейших (пропущенный через бактериальные фильтры и обработанный бактерицидами).
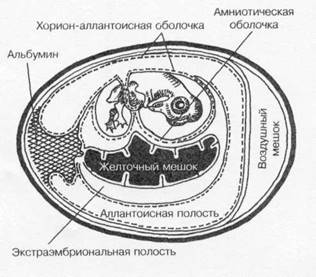 |
Заражение в амниотическую полость. Обычно используют 7-14-суточные эмбрионы, у которых после отслоения хорион-аллантоисной оболочки (см. выше) расширяют отверстие, захватывают пинцетом амниотическую оболочку и выводят через хорион-аллантоисную оболочку. Через неё в амниотическую полость вводят исследуемый материал.
Заражение в аллантоисную по лость. 10-суточные эмбрионы заражают через отверстия, сделанные в скорлупе и подлежащих оболочках (см. выше).
Заражение в желточный мешок. Используют 38-суточные эмбрионы, у которых в этом возрасте желточный мешок занимает почти всю полость яйца. Заражение проводят через отверстие, сделанное в воздушном мешке
Наблюдение и учёт результатов. В качестве вируссодержащего материала можно использовать содержимое желточного мешка, аллантоисную и амниотическую жидкости либо весь эмбрион, нарезанный вместе с окружающими
тканями на кусочки. Для выявления Рис. 1-20.Схематическое изображение
характерных поражений на хорион- развивающегося куриного эмбриона.
аллантоисной мембране удаляют скорлупу
и наружную оболочку. Затем мембрану извлекают и помещают в стерильную воду. Характер поражений изучают на тёмном фоне.
Животные модели
При невозможности выделить и идентифицировать вирус стандартными методами in vitro инфекционный материал вводят чувствительным к возбудителю животным, и после развития типичного инфекционного процесса проводят повторное заражение чувствительных клеточных культур. Наиболее часто используют мышей, кроликов и обезьян; для выделения некоторых вирусов (например, вирусов Коксаки) заражают мышат-сосунков. Вследствие дороговизны и сложности содержания лабораторных животных, практически повсеместно их вытеснили клеточные культуры. Тем не менее, животные модели активно используют для изучения особенностей патогенеза и формирования иммунных реакций при вирусных инфекциях.
Идентификация вирусов
Качественное определение
Наличие и биологическую активность вирусов определяют по эффектам, наблюдаемым на животных моделях (повышение температуры тела, появление характерных клинических признаков, гибель и т.д.), куриных эмбрионах и на клетках (в культурах). Под воздействием конкретных вирусов возможно изменение морфологии, роста, репродукции клеток либо их разрушение. Факт размножения вирусов в чувствительных клетках in vitro определяют по цитопатическим эффектам (в том числе бляшкообразованию, тельцам включений), феномену гемадсорбции, «цветной реакции».
Цитопатические эффекты оценивают при микроскопии клеточных культур. По степени поражения клеток выделяют вирусы с высокой или умеренной цитопатогенностью. Размножение вирусов в культурах клеток сопровождается нарушениями морфологии клеток монослоя. Некоторые вирусы вызывают характерные цитопатические изменения, что (с учётом клинической картины заболевания) позволяет быстро поставить предварительный диагноз. Например, размножение парамиксовирусов (вирусы кори, паротита, PC-вирус) сопровождается появлением характерных гигантских многоядерных клеток; аденовирусы вызывают образование скоплений больших круглых клеток, а при репродукции герпесвирусов клетки округлой формы диффузно располагаются по всему монослою.
Бляшкообразование. «Бляшками» называют негативные колонии — участки разрушенных клеток, выглядящие как зоны просветления на монослоях клеток, покрытых слоем агара. В некоторых случаях дозу и цитопатогенность вируса выражают в бляшкообразующих единицах (БОЕ).
Тельца включений. Многие вирусы вызывают появление в заражённых клетках характерных образований — скоплений вирусных белков или частиц, видимых в световой микроскоп. Тельца включений могут располагаться как в цитоплазме (тельца Гварнери при оспе), так и в ядрах клеток (аденовирусы).
Отсутствие цитопатического эффекта. Некоторые вирусы (например, вирус краснухи) не проявляют цитопатического эффекта. Их можно выявлять по интерференции другого вируса, способного вызывать дегенерацию заражённых клеток.
Феномен гемадсорбции. Многие заражённые вирусами клетки приобретают способность сорбировать на своей поверхности различные эритроциты. Феномен гемадсорбции имеет общие механизмы с гемагглютинацией и проявляется на ранних сроках, до проявления цитопатического эффекта, при его отсутствии либо слабой выраженности.
«Цветная реакция». В культуральную среду, используемую для поддержания клеток, вносят индикатор. Рост клеток сопровождается накоплением метаболитов, сдвигом рН среды и изменением окраски индикатора. Заражение культур вирусом резко ингибирует клеточный метаболизм, и среда сохраняет первоначальный цвет.
Экспресс-диагностика. Для быстрой идентификации вирусной инфекции разработаны многочисленные методы экспресс-диагностики, основанные на обнаружении вирусных Аг. Например,
Количественное определение
Количественное определение вирусов проводят двумя путями — изучением инфекционности и количественным определением вирусных Аг. Определение титра инфекционности вирусов в значительной степени зависит от метода количественного исследования; у бактериофагов отношение инфекционность-частица составляет приблизительно 1 (то есть каждая вирусная частица способна вызвать инфекцию), для вирусов животных данное отношение составляет 1:10 (иногда выше из-за вирусингибирующего действия факторов резистентности).
Определение инфекционности вирусов. Наиболее доступная форма количественного определения — подсчёт числа вирусных «бляшек». Прямые тесты на инфекционность применяют для установления инфекционной дозы (ID) или летальной дозы (LD) изучаемого вируса (обычно выражают в lg ). ID50 — разведение, инфицирующее 50% клеток; LD50 — разведение, убивающее 50% поражённых клеток или животных.
Выявление вирусных Аг и вирусных частиц. Наиболее распространённый метод — реакция количественной гемагглютинации. Метод основан на способности вирусов сорбироваться на поверхности эритроцитов животных и человека. Количественную электронную микроскопию применяют для подсчёта общего числа вирусных (но не инфекционных) частиц в исследуемом обьекте (например, культуральной жидкости).
Морфология вирусов
Изучение морфологии вирусов возможно лишь при помощи электронной микроскопии, однако чаще всего этот метод недоступен из-за отсутствия столь дорогого и сложного прибора. Более того, многие возбудители морфологически сходны, что снижает ценность этого метода. Наиболее распространён метод микроскопии содержимого везикул и тканевых экстрактов, обработанных красителями (негативное контрастирование), с последующим подсчётом ДНК- или РHK-содержащих вирусов. Электронная микроскопия позволяет быстро обнаружить орто- и парамиксовирусы в отделяемом дыхательных путей, герпесвирусы в жидкости везикул и ротавирусы в фекалиях.
Серологические методы идентификации
При большинстве вирусных инфекций развиваются иммунные реакции, применяемые для диагностики. Клеточные реакции обычно оценивают в тестах цитотоксичности лимфоцитов в отношении инфекционных агентов или заражённых ими клеток-мишеней либо определяют способность лимфоцитов отвечать на различные Аг и митогены. В работе практических лабораторий выраженность клеточных реакций определяют редко. Большее распространение нашли методы идентификации противовирусных AT.
РН основана на подавлении цитопатогенного эффекта после смешивания вируса со специфичными AT. Неизвестный вирус смешивают с известными коммерческими антисыворотками и после соответствующей инкубации вносят в монослой клеток. Отсутствие гибели клеток указывает на несоответствие инфекционного агента и известных AT .