Организм как открытая саморегулирующаяся система. Единство организма и внешней среды. Гомеостаз
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
Темновая адаптация происходит при переходе от больших яркостей к малым. Если глаз первоначально имел дело с большими яркостями, то работали колбочки, палочки же были ослеплены, родопсин выцвел, черный пигмент проник в сетчатку, заслоняя колбочки от света. Если внезапно яркость видимых поверхностей значительно уменьшится, то вначале раскроется шире отверстие зрачка, пропуская в глаз больший световой поток. Затем из сетчатки начнет уходить черный пигмент, родопсин будет восстанавливаться, и только когда его наберется достаточно, начнут функционировать палочки. Так как колбочки совсем не чувствительны к очень слабым яркостям, то сначала глаз не будет ничего различать, и только постепенно приходит в действие новый механизм зрения. Лишь через 50-60 мин пребывания в темноте чувствительность глаза достигает максимального значения. Световая адаптация – это процесс приспособления глаза при переходе от малых яркостей к большим. При этом происходит обратная серия явлений: раздражение палочек благодаря быстрому разложению родопсина чрезвычайно сильно, они «ослеплены», и даже колбочки, не защищенные еще зернами черного пигмента, раздражены слишком сильно. Только по истечении достаточного времени приспособление глаза к новым условиям заканчивается, прекращается неприятное чувство ослепления и глаз приобретает полное развитие всех зрительных функций. Световая адаптация продолжается 8-10 мин. Итак, адаптация обеспечивается тремя явлениями:
Зрачок может изменяться в диаметре от 2 до 8 мм, при этом его площадь и, соответственно, световой поток изменяются в 16 раз. Сокращение зрачка происходит за 5 сек, а его полное расширение – за 5 мин. Бинокулярное зрение (зрение двумя глазами) играет важную роль в восприятии разноудаленных предметов и определении расстояния до них, дает более выраженное ощущение глубины пространства по сравнению с монокулярным зрением, т.е. зрением одним глазом. При рассматривании предмета двумя глазами его изображение может попадать на симметричные (идентичные) точки сетчаток обоих глаз, возбуждения от которых объединяются в корковом конце анализатора в единое целое, давая при этом одно изображение. Если изображение предмета попадает на неидентичные (диспаратные) участки сетчатки, то возникает раздвоение изображения. Процесс зрительного анализа пространства зависит не только от наличия бинокулярного зрения, существенную роль в этом играют условно-рефлекторные взаимодействия, складывающиеся между зрительным и двигательным анализаторами. Определенное значение имеют конвергенционные движения глаз и процесс аккомодации, которые управляются по принципу обратных связей. Восприятие пространства в целом связано с определением пространственных отношений видимых предметов - их величины, формы, отношения друг к другу, что обеспечивается взаимодействием различных отделов анализатора; значительную роль при этом играет приобретенный опыт.
СЛУХ является результатом субъективного восприятия механической энергии колебаний воздуха. Его обеспечивает СЛУХОВОЙ АНАЛИЗАТОР. ОРГАН СЛУХА включает в себя:
Он состоит из 3 частей:
НАРУЖНОЕ УХО включает в себя:
СРЕДНЕЕ УХОвключает в себя цепь, соединённых между собой косточек: молоточек, наковальню и стремечко (связано через свое основание с овальным окном, а через него с внутренним ухом).
ВНУТРЕННЕЕ УХО содержит рецепторный аппарат слухового анализатора (улитка с кортиевым органом). Улитка - костная структура в виде спирали длиной около 35 мм, что составляет 2,5 завитка. Улитка разделена двумя мембранами (вестибулярной и основной) на три канала:
Верхний и нижний каналы связаны с помощью ГЕЛИКОТРЕМЫ у верхушки улитки и заканчиваются круглым окном.
Средний канал заполнен эндолимфой, которая по химическому составу приближается к внутриклеточной жидкости (высокое содержание калия). Средний канал содержит (на основной мембране) рецепторный аппарат – КОРТИЕВ ОРГАН, который образован механорецепторами (содержат 4 ряда ВОЛОСКОВЫХ клеток). Они прикрыты ТЕКТОРИАЛЬНОЙ (покровной) мембраной, которая имеет свободный край и при передаче звука сгибает волоски рецепторных клеток. Это преобразует акустические сигналы в потенциалы нервной системы. Преобразование звуковых сигналов в электрические:
Ухо человека может-воспринимать звук при колебании воздуха в диапазоне от 16 до 20 000 Гц. Высказывают предположение, что есть два механизма различения тонов. Звуковая волна, создаваемая колебанием молекул воздуха, распространяется в виде продольной волны давления. Передаваясь на перилимфе и эндолимфе, она между пунктами возникновения и затухания имеет участок с максимальной амплитудой колебаний. Место расположения этого участка зависит от частоты колебаний: при высоких частотах она лежит ближе к овальной мембраны, а при низких - ближе к геликотремы. Вследствие этого амплитудный максимум для каждой частоты проявляется в специфической точке эндолимфатического канала. Расположенные здесь сенсорные клетки возбуждаются сильнее. В этом заключается так называемая пространственная теория кодирования высоты тона, который воспринимается в самом рецепторе. Кроме того, считают, что при небольшой частоте колебаний (до 1000 Гц) может действовать телефонная принцип кодирования: потенциал действия в кохлеарном нерве возникает с частотой, которая является резонансным до частоты звуковых колебаний. В рецепторах только начинается различение звуковой информации. Обработка завершается в нервных центрах.
Проведение возбуждения в нервные центры: Осуществляется через Спиральный ганглий улитки, где расположены нейроны первого порядка. Его отростки образуют Слуховой или кохлеарный нерв, который направляется в Кохлеарные ядра продолговатого мозга, где расположены нейроны второго порядка. По их отросткам возбуждение направляется к Верхней оливе, где происходит первый перекрёст слуховых путей. Далее возбуждение поступает в Задние бугры четверохолмия (второй перекрёст слуховых путей), к Внутренним коленчатым телам и Слуховой коре, которая расположена в верхней части височной доли и где происходит третий перекрёст слуховых путей. ОТДЕЛЬНЫЕ ЧАСТИ СЛУХОВОЙ ПРОВОДЯЩЕЙ СИСТЕМЫ обеспечивают определённые ФУНКЦИИ
Кохлеарный нерв достигает вентрального и дорсального кохлеарных ядер. Волокна от вентрального ядра направляются как к ипси-, так и в контралатеральный оливарних комплексов. Дорсальный кохлеарный тракт переходит на противоположную сторону и заканчивается в ядре латеральной петли. Нейроны, которые поднимаются из масел, также отдают коллатерали ядрам латеральной петли. Далее волокна идут в нижних холмиков чотиригорбкового тела и медиального коленчатого тела. Затем они заходят в ме- таталамус, и только после этого звуковые пути попадают в первичной звуковой зоны коры. Рядом с ней находятся нейроны, которые относятся к вторичной звуковой зоны коры большого мозга. Информация, содержащаяся в звуковом стимуле, проходя через названные ядра переключения, много раз (как правило, не менее 5-6 раз) переписывается в виде нейронного возбуждения. При этом на каждом этапе она анализируется, причем нередко с подключением сенсорных сигналов других («неслухових») отделов ЦНС. Вследствие этого могут возникнуть рефлекторные ответы, характерные для определенного отдела ЦНС. Но только в коре большого мозга появляется ощущение определенного звука. Нейроны вентрального ядра еще воспринимают чистые тона, т.е. возбуждения в них возникает при воздействии строго определенных тонов. В дорсальном же ядре лишь незначительная часть нейронов возбуждается чистыми тонами. Другие нейроны реагируют на сложный стимул, например, на изменение частоты, снижение звука и т.д.. На высших уровнях в отдельных нейронах постепенно усиливается специфичность реагирования на сложные звуковые модуляции. Так, одни нейроны возбуждаются только при изменении амплитуды звука, другие - изменении частоты, третьи - при варьировании расстояния от источника, его перемещении. Таким образом, каждый раз при действии реально существующих в природе сложных звуков в нервных центрах возникает своеобразная мозаика возбуждения нейронов. Происходит запоминание этой мозаичной карты, обусловленной поступлением соответствующего звука. Люди могут оценивать различные свойства звука только при соответствующей тренировке. Корковые нейроны активизируются по-разному: одни - контралатеральный ухом, другие - ипсилатерально стимулами, третьи - только при одновременной стимуляции обоих ушей. Возбуждаются они, как правило, целыми звуковыми группами. Повреждение этих отделов ЦНС ухудшает восприятие речи, пространственную локализацию источника звука.
ВНУТРЕННЕЕ УХО содержит рецепторный аппарат вестибулярного анализатора (преддверие и полукружные каналы). Вестибулярный анализатор анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения тела, а также при изменении положения головы в пространстве. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры. Среди вестибулярных реакций на первом месте находятся статистические и статокинетические реакции, обеспечивающие сохранение равновесия при изменении положения тела и его частей или при возникающих ускорениях во время перемещения тела в пространстве. В осуществлении этих реакций участвуют также и проприорецепторы мышц. Рецепторы статолитовых органов и полукружных каналов: Вестибулярный орган состоит из статолитового аппарата и трех полукружных каналов, расположенных во внутреннем ухе в трех взаимно перпендикулярных плоскостях: фронтальной, сагиттальной и горизонтальной. Возбуждающим фактором для вестибулорецепторов, представленных волосковыми клетками, является наклон волосков вследствие смещения отолитовой мембраны при линейных ускорениях. Рецепторные клетки, находящиеся в ампулах, возбуждаются при угловых ускорениях вследствие движения эндолимфы по полукружным каналам. Вестибулорецепторы относятся к вторичночувствующим и связаны через синапсы с афферентными волокнами нейронов вестибулярного ганглия, расположенного в височной кости. Вестибулярные ганглии и ядра: От вестибулярных ганглиев волокна вестибулярного нерва направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, поступают к нейронам бульбарного вестибулярного комплекса: предверное верхнее ядро Бехтерева, предверное латеральное ядро Дейтерса, ядро Швальбе. Из вестибулярных ядер возбуждения направляются по вестибулоспинальному тракту к мотонейронам мышц-разгибателей; непосредственно к мотонейронам шейного отдела спинного мозга; к глазодвигательным ядрам и мозжечку; к ретикулярной формации и через таламус к задней центральной извилине коры большого мозга. Функциональные связи между вышеуказанными структурами обеспечивают не только поддержание позы человека (сохранение равновесия), но и координацию двигательных актов при выполнении целенаправленной деятельности.
БОЛЬ является интегративной функцией организма, которая мобилизует организм и его разнообразные функциональные системы на защиту от воздействующих вредящих факторов и включает такие компоненты, как сознание, ощущение, память, мотивации, вегетативные, соматические, поведенческие реакции, эмоции. При этом внешние или внутренние повреждающие воздействия изменяют НОРМАЛЬНУЮ ЖИЗНЕДЕЯТЕЛЬНОСТЬ ОРГАНОВ и ТКАНЕЙ организма. Возникающее раздражение ноцицепторов вызывает афферентную импульсацию к различным структурам ЦНС, где формируется болевое ощущение. Следствием являются эффекторные влияния, направленные на устранение вредоносного фактора, щажение больного органа, компенсаторную мобилизацию защитных сил организма. I. Классификация боли по эволюционному механизму
II. Классификация боли по месту возникновения
III. Классификация боли по времени формирования
IV. Особые формы боли
ОЩУЩЕНИЕ боли является отрицательной биологической потребностью организма, связанной с нарушением целостности защитных покровных оболочек и изменением уровня кислородного дыхания тканей БОЛЕВЫЕ рецепторы или НОЦИЦЕПТОРЫ являются высокопороговыми рецепторами. Они представляют свободные окончания немиелинизированных волокон, образующие плексиморфные сплетения в тканях кожи, мышц и некоторых органов Ноцицепторы подразделяются на МЕХАНОНОЦИЦЕПТОРЫ и ХЕМОНОЦИЦЕПТОРЫ, которые возбуждаются при воздействии сильных повреждающих раздражителей в результате механического смещения мембраны или действия химических веществ.
Механоноцицепторы обеспечивают сохранность защитных оболочек организма, изолирующих внутреннюю среду от внешнего мира, и реагируют на уколы, сжатие, скручивание, давление, сгибание, температуру. Хемоноцицепторы обеспечивают контроль тканевого дыхания и реагируют на повреждение тканей, развитие воспаления (нарушение метаболизма, сопровождающееся выделением гистамина, простагландинов, хининов, всех веществ, подавляющих окислительные процессы), а также на прекращение доступа кислорода к тканям (ишемия). АФФЕРЕНТНЫЕ НОЦИЦЕПТИВНЫЕ ВОЛОКНА включают:
Их активация формирует первую боль
Их активация формирует вторую боль и тонические сокращения мышц. Возбуждение по ним поступает в ЗАДНИЕ РОГА СПИННОГО МОЗГА, средний мозг, Гипоталамус, Таламус, Лимбические структуры переднего мозга, сенсорные и Ассоциативные зоны коры. Компоненты системной болевой реакции:
Механизмы контроля болевой чувствительности:
ОПИАТНЫЕ ПЕПТИДЫ – это эндогенные морфиноподобные вещества, которые вырабатываются в гипоталамусе и гипофизе. Их представителями являются: ЭНДОРФИНЫ и ЭНКЕФАЛИНЫ. Антагонистом является НАЛОКСОН (блокирует опиатные пептиды). При БОЛИ их содержание СНИЖАЕТСЯ. При АНАЛЬГЕЗИИ содержание УВЕЛИЧИВАЕТСЯ. Количество опиатных РЕЦЕПТОРОВ и опиатных ПЕПТИДОВ определяет порог БОЛЕВОЙ ЧУВСТВИТЕЛЬНОСТИ (понижение опиатных пептидов вызывает повышение болевой чувствительности – состояние ГИПЕРАЛГЕЗИИ).
Серотонин выделяется некоторыми нейронами ствола мозга, которые оказывают нисходящие влияния на пути болевой чувствительности. При БОЛИ выделение серотонина УМЕНЬШАЕТСЯ. При АНАЛЬГЕЗИИ его содержание УВЕЛИЧИВАЕТСЯ. УМЕНЬШЕНИЕ выделения серотонина ПОВЫШАЕТ болевую чувствительность.
Прямые проекции от гипоталамуса к нейронам заднего рога спинного мозга имеют катехоламиновую природу. Катехоламины в большой концентрации УГНЕТАЮТ ноцицептивную импульсацию. При отсутствии болевого раздражителя НОЦИЦЕПТИВНАЯ И АНТИНОЦИЦЕПТИВНАЯ СИСТЕМЫ находятся в равновесии. НОЦИЦЕПТИВНАЯ СИСТЕМА формирует болевое ощущение. АНТИНОЦИЦЕПТИВНАЯ СИСТЕМА подавляет болевое ощущение, тормозит активность ноцицептивной системы и определяет ПОРОГИ возбудимости НОЦИЦЕПТОРОВ. К НОЦИЦЕПТИВНЫМ СТРУКТУРАМ относятся задние рога спинного мозга, таламус. Они продуцируют НОЦИЦЕПТИВНЫЕ ВЕЩЕСТВА: вещество «Р», брадикинин, гистамин, соматостатин. К АНТИНОЦИЦЕПТИВНЫМ СТРУКТУРАМ относятся: центральное серое околоводопроводное вещество, ядра шва, дорсомедиальный гипоталамус. Там выделяются АНТИНОЦИЦЕПТИВНЫЕ ВЕЩЕСТВА: катехоламины, эндорфины, энкефалины, серотонин, ацетилхолин, окситоцин, глицин, нейротензин. НОЦИЦЕПТИВНЫЙ РАЗДРАЖИТЕЛЬ вызывает торможение АНТИНОЦИЦЕПТИВНОЙ СИСТЕМЫ и активацию НОЦИЦЕПТИВНОЙ СИСТЕМЫ. Следствием является БОЛЕВОЕ ОЩУЩЕНИЕ.
Принципиально выделяется два пути обезболивания: снижение активности ноцицептивной системы и повышение активности антиноцицептивной системы. Это достигается при помощи:
Фармакологические меры
Нейрохирургические меры – хирургическое прекращение поступления ноцицептивных сигналов (хордотомия). Из-за необратимости этих мер применяют только при хронических болях, доставляющих мучения человеку; Психогенная регуляция болевых ощущений предусматривает корковую регуляцию болевой чувствительности и изменение эмоционального состояния. Так, предупреждение человека о воздействии болевого раздражителя, гипноз и внушение снижают болевую чувствительность. Положительные эмоции оказывают антиноцицептивное влияние. К нетрадиционным методам обезболивания относятся:
Система кpови пpедставляет собой упоpядоченную взаимосвязь элементов, обладающих собственной оpганизацией, стpуктуpой и pегуляцией. Система крови включает в себя следующие компоненты: Кpовь, циpкулиpующая в сосудах (пpедставлена фоpменными элементами и жидкой частью) Аппаpат кpоветвоpения, котоpый включает оpганы, пpодуциpующие фоpменные элементы (костный мозг, лимфостpуктуpы) и оpганы, пpодуциpующие элементы жидкой части (печень и дp.) Аппаpат кpовеpазpушения (печень, селезёнка, костный мозг) Аппаpат депониpования кpови (синусы костного мозга, лимфоузлы, печень, селезёнка, стенки сосудов лёгких и кожи) Аппаpат pегуляции, включающий нейpо-гоpмонально-гумоpальные механизмы, ответственные за обеспечение оpганизма кpовью, адекватной его потpебностям Основными функциями кpови являются : 1. Тpнаспоpтная (тpанспоpт pазличных веществ в пpеделах оpганизма) 2. Питательная (кpовь пpиносит клеткам питательные вещества) 3. Дыхательная (обеспечивает газообмен О2 и СО2) 4. Регулятоpная (обеспечивает гомеостаз, гумоpальную pегуляцию) 5. Выделительная (доставляет пpодукты pаспада к оpганам выделения) 6. Теpмоpегулятоpная (поддеpжание темпеpатуpы тела за счёт изменения теплоотдачи) 7. Защитная (адсоpбция токсических веществ, фагоцитоз, обpазование антител, иммуннитет, гемостаз) Кpовь состоит из фоpменных элементов (45 %) и жидкой части или плазмы (55 %) Фоpменные элементы включают эpитpоциты, лейкоциты, тpомбоциты
Общее количество кpови в оpганизме взpослого человека составляет 6-8 % от массы тела (пpи массе 70 кг – это 5-6 л), из котоpой около половины циpкулиpует, а остальная часть находится в депо (в печени – 20 %, в селезёнке – до 16 %, в кожных сосудах – до 10 %) Гематокрит — это соотношение объёмов плазмы крови и форменных элементов. 1. Соотношение определяется путём центрифугирования крови в специальном капилляре с делениями — гематокрите. 2. В нормальных условиях это соотношение составляет 45 % форменных элементов и 55 % плазмы. 3. Эта величина у здорового человека может претерпевать существенные и достаточно длительные изменения лишь при адаптации к большим высотам.
Плазма крови - жидкая часть крови, остающаяся после удаления ее форменных элементов. Плазма, лишенная фибриногена, называется сывороткой. В состав плазмы входят: 1) вода (90-92 %) 2) сухой остаток (8-10%) Сухой остаток состоит из: 1) оpганических веществ 2) неоpганических веществ К оpганическим веществам крови относятся : 1. Белки плазмы (общее количество 7-8 %) – альбумины (4,5 %), глобулины (2-3,5 %), фибpиноген (0,2-0,4 %) 2. Hебелковые азотсодеpжащие соединения (аминокислоты, полипептиды, мочевина, мочевая кислота, кpеатин, кpеатинин, аммиак) Общее количество небелкового азота (остаточный азот) составляет 11-15 ммоль/л (30-40 мг%). Пpи наpушении функции почек, выделяющих шлаки из оpганизма, содеpжание остаточного азота pезко возpастает 3. Безазотистые оpганические вещества: глюкоза 4,4-6,65 ммоль/л (80-120 мг%), нейтpальные жиpы, липиды 4. Феpменты и пpофеpменты: некотоpые из них участвуют в пpоцессах свёpтывания кpови и фибpинолиза (пpотpомбин, пpофибpинолизин), некотоpые - pасщепляют гликоген, жиpы, белки и дp. Hеоpганические вещества плазмы составляют около 1% от её состава К ним относятся пpеимущественно катионы (Na+, Ca2+, K+, Mg2+) и анионы (Cl-, HPO42-, HCO3-) Из тканей оpганизма в кpовь поступает большое количество пpодуктов обмена, биологически активных веществ (сеpотонин, гистамин), гоpмонов, из кишечника всасываются питательные вещества, витамины Однако состав плазмы от этого существенно не изменяется. Постоянство состава плазмы обеспечивают pегулятоpные механизмы, восстанавливающие состав и свойства внутpенней сpеды
Электролитный состав плазмы важен для поддержания ее осмо тического давления, кислотно-щелочного состояния, функций кле точных элементов крови и сосудистой стенки, активности фермен тов, процессов свертывания крови и фибринолиза. Поскольку плаз ма крови постоянно обменивается электролитами с микросредой клеток, содержание в ней электролитов в значительной мере опре деляет и фундаментальные свойства клеточных элементов органов — возбудимость и сократимость, секреторную активность и проница емость мембран, биоэнергетические процессы. Содержание основных электролитов в плазме крови, эритроцитах и тканевой микросреде представлено в табл.2.1. 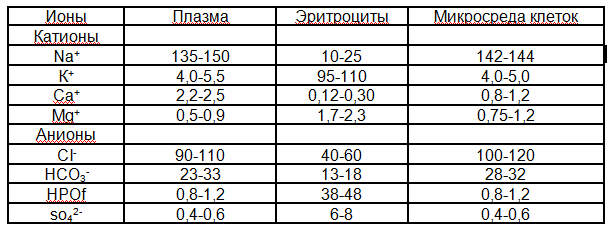 Таблица 2.1. Содержание электролитов в плазме крови, эритроцитах и микросреде тканей (ммоль/л) у человека Из таблицы видно, что содержание натрия и калия в плазме и эритроцитах отличается также, как и в других клетках и внеклеточной среде (глава 1), и, соответственно, обуслов лено различиями проницаемости мембран и работой К- Na- насосов клеток. Часть катионов плазмы связана с анионами органических кислот и белков, что играет роль в поддержании кислотно-щелоч ного состояния и необходимо для реализации функций белков. Отличается в плазме и эритроцитах содержание и ряда анионов, прежде всего хлора и бикарбоната. Эти различия обусловлены об меном этих анионов между эритроцитами и плазмой в капиллярах легких и тканей при дыхании. Содержание натрия и калия в плазме крови — жесткие гомеостатические константы, зависящие от баланса процессов поступления и выведения ионов, а также их перераспределения между клетками и внеклеточной средой. Регуляция гомеостазиса этих катионов осу ществляется изменениями поведения (большее или меньшее потреб ление соли) и системами гуморальной регуляции (см.главу 3), среди которых основное значение имеют ренин-ангиотензин-альдостероновая система и натриуретический гормон предсердий (см.главу 5). Жесткой гомеостатической константой является и концентрация кальция в плазме крови. Кальций содержится в двух формах: свя занной (с белками, в комплексных соединениях, малорастворимых солях) и свободной, ионизированной (Са++). Основные биологичес кие эффекты кальция обусловлены его ионизированной формой. В цитозоле клеток ионизированного кальция содержится мало, но его количество чрезвычайно тонко регулируется, поскольку этот катион является важнейшим регулятором обменных процессов и функций клеток. Поступление кальция в клетки из внеклеточной среды свя зано с его уровнем в микросреде и плазме крови, хотя в большей степени зависит от специальных транспортных мембранных меха низмов (каналов, насосов, переносчиков). В клеточном цитозоле ионизированный кальций связывается с белками, а также удаляется с помощью специальных Са-насосов во внутриклеточные депо (ми тохондрии, цитоплазматический ретикулум) и наружу в микросреду клеток. Содержащийся в плазме крови ионизированный кальций помимо того, что является источником для транспорта внутрь кле ток, необходим для обеспечения физико-химических свойств плаз менных белков, активности ферментов, например, для реализации механизмов свертывания крови. Регуляция уровня ионизированного кальция в плазме крови осуществляется специальной гуморальной системой, включающей ряд кальций-регулирующих гормонов: око лощитовидных желез (паратирин), щитовидной железы (кальцитонин и его аналоги), почек (кальцитриол). В плазме крови содержится и большое число различных микро элементов, называемых так из-за очень малых концентраций. Как минимум 15 микроэлементов, содержащихся в плазме крови, напри мер, медь, кобальт, марганец, цинк, хром, стронций и др., играют важную роль в процессах метаболизма клеток и обеспечении их функций, поскольку входят в состав ферментов, катализируют их действие, участвуют в процессах образования клеток крови и гемог лобина (гемопоэзе) и др. Осмотическое давление крови равняется 7,6 атм. Зависит от концентрации в плазме крови электролитов неэлекролитов. На долю неорганических электролитов приходится до 96 %. Жидкость или раствор с осмотическим давлением 7,6 атм. называют изотоническим; более 7,6 атм. – гипертоническим, менее 7,6 атм. – гипотоническим. Под осмотической резистентностью эритроцитов понимается устойчивость эритроцитов по отношению к гипотоническим растворам натрия хлорида. Изотоническим (т. е. раствором, осмотическое давление которого равно осмотическому давлению в сыворотке крови) является 0,85% раствор натрия хлорида. Растворы его с меньшей концентрацией являются гипотоническими. Различают минимальную и максимальную резистентность эритроцитов:
У здоровых людей минимальная резистентность эритроцитов равна 0,45—0,50%, максимальная — 0,35—0,40% раствора натрия хлорида. Наименее устойчивы к гипотоническим растворам сфероциты. Наиболее выраженное понижение осмотической стойкости эритроци тов наблюдается при врожденной гемолитической анемии. Однако необходимо помнить, что и при приобретенной аутоиммунной гемоли тической анемии осмотическая стойкость эритроцитов иногда может быть снижена. Незначительное понижение осмотической стойкости эритроцитов может иногда наблюдаться при полицитемии, туберкулезе, лимфограну лематозе, циррозах печени, лейкозах. Повышение осмотической стойкости эритроцитов возможно при механической желтухе (что в сочетании с другими видами обследования помогает при дифференциальной диагностике этого вида желтухи с желтухой при врожденной гемолитической анемии), при талассемии и гемоглобинозах. Постоянство осмотического давления крови поддерживается главным образом благодаря деятельности почек, через которые выводится избыток воды и растворимых солей.
Белки плазмы составляют основную массу органических веществ плазмы крови. Белки плазмы крови подразделяются на: 1) альбумины (60,5 % от общего количества белков) 2) глобулины (35,4 %) 3) фибриноген (4,1 %) |