реферат пчеловодтво. реферат пчеловодство. Пчеловодство
 Скачать 0.55 Mb. Скачать 0.55 Mb.
|

|
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ ЗАОЧНЫЙ УНИВЕРСИТЕТ ФАКУЛЬТЕТ Зоотехния__(кинология и фелинология) 2 курс Коломна Реферат по теме пчеловодство. Зрение пчел и визуальная ориентация. ВЫПОЛНИЛ: СТУДЕНТ БАКАЛАВРИАТА Щербакова Анна Ильинична ГРУППЫ __________________ шифр №_044б____ ПРОВЕРИЛ: _____________________________ БАЛАШИХА – 2021 Зрение пчел и визуальная ориентация. 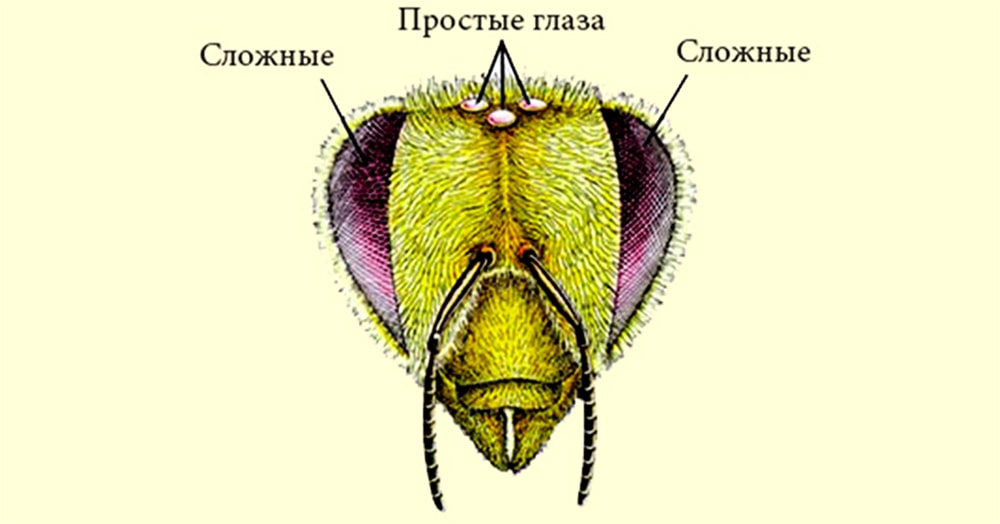 Фоторецепторы пчел представлены двумя сложными (фасеточными) и тремя простыми (дорсальными) глазами. Они существенно различаются по морфологическим и функциональным свойствам. Фасеточные глаза. Структурной единицей фасеточного глаза является омматидий. От их числа зависит площадь, занимаемая глазом. Число омматидиев в глазу рабочей пчелы колеблется от 4 тыс. до 5 тыс., у матки — от 3 тыс. до 4 тыс., а у трутня - от 7 тыс. до 9 тыс. Светопреломляющий аппарат, омматидия состоит из двух основных элементов: хрусталика и кристаллического, или хрустального, конуса. Хрусталик выполняет роль собирающей линзы, имеющей форму выпуклого шестигранника. Под ними находится кристаллический конус. Он образован четырьмя клетками (земперовыми, кристаллическими), скомпонованными в прозрачное тело грушевидной формы. Основание кристаллического конуса соприкасается с хрусталиком, а вершина подходит к ретинальным клеткам. В каждом омматидии имеется по восемь ретинальных (зрительных) клеток, сгруппированных тесным цилиндрическим пучком. Внутри него вдоль оптической оси омматидия расположен рабдом замкнутого (спаянного) типа, образованный рабдомерами ретинальных клеток. Каждый рабдомер состоит из большого количества микровилл — микроскопических плотно уложенных трубочек диаметром от 400 до 1200 ?. Микровиллы восьми ретинальных клеток рабдома расположены в двух взаимно перпендикулярных направлениях. В стенках микровилл находится зрительный пигмент, реагирующий на свет. Пигмент состоит из белка опсина и красящего вещества ретиналя, представляющего собой альдегид витамина А. Ретинальные клетки и кристаллический конус омматидия оптически изолированы от смежных омматидиев. Оптическим изолятором служит чехол из пигментных (ирисовых) клеток. Они препятствуют проникновению лучей света к рабдомам через фасетки соседних омматидиев и изолируют рабдомы от рассеяния света внутри глаза. Все это способствует повышению дирекциональной чувствительности омматидия. Разные ретинальные клетки омматидия обладают различной спектральной и поляризационной чувствительностью. Внешне омматидии, находящиеся в различных частях глаза, не имеют явно выраженных различий, хотя не все они функционально равноценны. В частности, анализ поляризации света возможен только в омматидиях, расположенных в верхней половине глаза, а различные формы и размеры объектов осуществляются омматидиями нижней половины глаза. Равноценными свойствами обладают омматидии любой части глаза в отношении различия цвета (Г. Мазохин — Поршняков, 1965, 1977). Механизм цветоразличия фасеточным глазом пчелы основан на наличии в каждом омматидии четырех светоприемников (светочувствительных веществ) с различной спектральной чувствительностью. Первый максимально чувствителен к свету с длиной волны 340 нм, второй — 430, третий — 460 и четвертый — 530 нм. Восприятие направления поляризации света, по-видимому, связано с поляризационными свойствами рабдомеров зрительных клеток. Важная роль в этом отводится взаимно перпендикулярному расположению микровилл, что приводит к различному возбуждению симметричных пар клеток омматидия в зависимости от плоскости, в которой колеблются проникающие в омматидии лучи света. Наличие же у каждой зрительной клетки собственного нервного волокна, связывающего ее с центральными отделами мозга, обеспечивает дифференциацию информации об уровне раздражения разных зрительных клеток омматидия. Дорсальные глазки.  Они расположены в виде треугольника на лобно-теменной поверхности головы. Диоптрический аппарат дорсальных глазков представлен хрусталиком, который является слоистым утолщением кутикулярного покрова. Под хрусталиком находится слой корнеагенных клеток, образовавших хрусталик. К слою корнеагенных клеток примыкает вогнутая по форме хрусталика сетчатка, сложенная из отдельных ретинул, которые группируются из удлиненных зрительных клеток. В дистальном конце каждой ретинулы, находится рабдом, который складывается из рабдомеров входящих в нее зрительных клеток. Проксимальный конец зрительной клетки переходит в нервное волокно, связывающее глазок с осцелярным ганглием. Они расположены в виде треугольника на лобно-теменной поверхности головы. Диоптрический аппарат дорсальных глазков представлен хрусталиком, который является слоистым утолщением кутикулярного покрова. Под хрусталиком находится слой корнеагенных клеток, образовавших хрусталик. К слою корнеагенных клеток примыкает вогнутая по форме хрусталика сетчатка, сложенная из отдельных ретинул, которые группируются из удлиненных зрительных клеток. В дистальном конце каждой ретинулы, находится рабдом, который складывается из рабдомеров входящих в нее зрительных клеток. Проксимальный конец зрительной клетки переходит в нервное волокно, связывающее глазок с осцелярным ганглием.Фокальная плоскость хрусталика глазка лежит далеко позади слоя зрительных клеток, в силу этого (хотя выделенный хрусталик и дает четкое изображение) на слое рабдомов получается размытое изображение. Это ставит под сомнение возможность восприятия деталей предметов с помощью дорсальных глазков. Электрофизические исследования показали, что форма электроответа изменяется при затенении и повторном освещении дорсальных глазков пчелы. Форма электро-ретинограммы при этом зависит не только от интенсивности света, но и от его длины волны (спектра). Спектральная чувствительность глазков характеризуется кривой с двумя вершинами: при длине волны 490 и 340 нм. Это указывает на присутствие двух цветовых приемников, то есть способность дорсальных глазков различать цвет. Они обладают также механизмом восприятия плоскости поляризации света. Закрашивание дорсальных глазков рабочих пчел укорачивает период их летной активности в течение светового дня: они позже начинают вылетать из улья и раньше прекращают в конце дня летную деятельность. Значит, дорсальные глазки служат для усиления фототаксических реакций. Визуальная ориентация. Глаза пчел, как и других насекомых, не обладают аккомодацией. Острота их зрения определяется угловой плотностью омматидиев. Поэтому пчела различает две детали, например светящиеся точки, если их проекция в фасеточном глазу захватывает не менее трех омматидиев. Отсюда понятна причина крайней близорукости пчелы. Она может различать тем больше деталей объекта, чем ближе от него находится. При движении пчелы на точечный источник света соблюдаемая ею точность направления лимитируется углом между осями смежных омматидиев. Если источник света, к которому летит пчела, сместить на угол, меньший указанного, то она не изменит направления полета. Для реализации процесса зрительного восприятия необходимо, чтобы ретинальное изображение перемещалось. Пчелиные глаза неподвижно закреплены на голове. Ретинальное изображение в них может перемещаться в случае движения рассматриваемого объекта или самой пчелы. Некоторое перемещение ретинального изображения возможно за счет колебаний тела, вызываемых работой дыхательной системы и сердца. Но это, по-видимому, не имеет существенного значения для визуальной ориентации пчел. Судя по поведению, они быстро обнаруживают и опознают предметы в то время, когда летают. Сидящая пчела четко реагирует на движущийся предмет. Способность пчел обнаруживать предметы и различать их детали основана на происходящем одновременно неодинаковом возбуждении разных омматидиев глаза. Предельная возможность различения летящей пчелой различных деталей предметов, попадающих в поле её зрения, лимитируется скоростью полета, так как с ней связана частота смены ретинальных изображений. Различать в полете быстро сменяющиеся детали предметов пчела может благодаря очень низкой инерционности зрительного аппарата. В электрофизиологических и поведенческих экспериментах (по оптомоторной реакции) установлено, что у пчел мелькания не сливаются до тех пор, пока их частота не превосходит 250—300 Гц. Это более чем на порядок выше возможностей человеческого глаза. Он не замечает мельканий кадров изображений, следующих с частотой 16 Гц, что используется в кинотелевизионной технике. До недавнего времени считалось, что пчелы четко дифференцируют различные геометрические фигуры исключительно по степени их расчлененности, но не могут различать их форму. Это мнение было опровергнуто оригинальными опытами Г. Мазохина-Поршнякова и Т. Вишневской. Исследователи приучали пчел посещать составные фигуры. Каждая из них, например треугольник, был набран из большого количества маленьких треугольников, крут — из маленьких кружков и т. п. В результате пчелы, получавшие корм на одной из таких фигур, в дальнейшем подлетали в первую очередь к этой фигуре, хотя ее местоположение постоянно меняли. На основании этих опытов было доказано, что пчелы могут различать треугольник, круг и квадрат. Более того, пчелы способны отличать одинаковые фигуры по размеру. Точность различия размера возрастает с увеличением площади фигуры и степени ее расчлененности. К. Фриш еще в 1914 г. доказал способность пчел ориентироваться, используя цветные ориентиры. Для этого пчел вначале приучали к подкормке на квадрате из синей бумаги. Она находилась среди 15 других, но ахроматических (серых), отличающихся по степени яркости от белой до черной. Бумажные квадраты размещали в шахматном порядке. В процессе опытов положение синего и других квадратов произвольно меняли. Несмотря на это, пчелы находили синий квадрат среди других даже в тех случаях, когда на нем не было подкормки. Способность пчел использовать цветовые ориентиры была подтверждена многочисленными работами отечественных и зарубежных исследователей. Сравнительно недавно Г. Мазохиным-Поршняковым (1969), изучавшим возможности цветовой ориентации насекомых, была выявлена способность пчел различать близкие окраски, например такие, как желтую и оранжевую, а желто-оранжевые тона от зеленых. Этим дан, наконец, ответ на вопрос о том, как пчелы находят желтые цветы на фоне зеленых листьев. Механизм различия цветов основан, по-видимому, на использовании глазом пчелы спектральных отражательных характеристик освещенных поверхностей. Они в зависимости от цвета освещения отражают свет разного спектрального состава. Его различие основано на константности цветовосприятия, что позволяет дифференцировать предметы в разных условиях освещения.  Важная особенность различия цветов пчел связана с тем, что их коротковолновый цветовой приемник омматидия воспринимает ультрафиолет. Благодаря этому пчелы дифференцируют отражения разных поверхностей в ультрафиолете. Например, пчелы отличают одинаковые (белые) для глаза человека поверхности, окрашенные цинковыми и свинцовыми белилами. Дело здесь в том, что свинцовые белила сильнее, чем цинковые, отражают ультрафиолетовые лучи. Поэтому первые воспринимаются пчелами как «действительно белые», а вторые — дополнительно с сине-зеленым цветом. Из-за отражения лучей разного спектрального состава венчиками цветов пчелы видят их не такими, как человек. Так, цветок красного мака отражает и красные, и ультрафиолетовые лучи. Но, так как глаз пчелы не воспринимает красного цвета, то она видит венчик мака чисто ультрафиолетовым. У некоторых растений только части цветков отражают ультрафиолет. В таких случаях цветки имеют для глаза пчел дополнительные (скрытые для человеческого глаза) рисунки. Например, периферическая часть венчика цветка гусиной лапки отражает ультрафиолетовые лучи сильнее, чем центральная. Поэтому он воспринимается глазом пчелы как двуцветный. Скрытый рисунок цветков служит пчелам дополнительным визуальным ориентиром и является указанием местоположения в цветке нектара и пыльцы. Пользуясь им, пчела располагает дополнительным средством для опознавания нужных цветков.  Все изложенное выше указывает на то, что пчелы хорошо различают и запоминают предметы по цвету и форме. Но их легко дезориентировать, переставив улей со своего места на небольшое расстояние. Пчел особенно дезориентирует, если рядом с ульем перемещаемой семьи находятся другие ульи, а также предметы, которые могут служить ориентирами. В таких случаях возвращающиеся пчелы долго кружатся («зависают») у того места, где находилось летковое отверстие их улья. Если же в период высокой летной активности, когда пчелы интенсивно пополняют пищевые запасы, на место улья с отставленной семьей поставить пустой, то в нем собираются через некоторое время практически все пчелы, занимающиеся доставкой корма. Такая картина наблюдается, если улей с пчелами отставить всего на 1,5—3 м. Чем же объяснить, что пчелы, так хорошо различающие оттенки и детали предметов, не летят в свой улей, находящийся в поле их зрения? Дело в том, что пчелы запоминают не только детали своего жилища, но место его положения относительно деревьев и других ориентиров в течение первых вылетов из улья. Для этого они совершают круговые облеты. В дальнейшем пчелы летят к цели без предварительных ориентировочных облетов. Точность подлета пчел к месту, где находился их улей, говорит о хорошей ориентационной способности и памяти пчел. Подтверждением того, что в первые вылеты пчелы запоминают место положения своего улья, служит следующее. Перемещение улья в то время, когда пчелы не летают, например, в период летних похолоданий, не так сильно дезориентирует пчел впоследствии (с наступлением хорошей погоды и возобновлением летной деятельности). К старому месту их подлетает тем меньше, чем дольше пчелы не вылетали из улья. Именно с этим прямо связано количество особей, совершающих облеты, прежде чем отправиться на поиски и доставку корма. При низкой летной активности, когда пчелы слабо пополняют свои запасы, вылетая из улья, они чаще совершают облеты (кружатся над ним, летают по зигзагообразной траектории, прежде чем улетать). В то время, когда идет интенсивное пополнение корма, большая часть пчел не совершает облетов. Они не задерживаются ни у леткового отверстия, ни после взлета. Поднявшись в воздух, сразу же отправляются к цели полета. 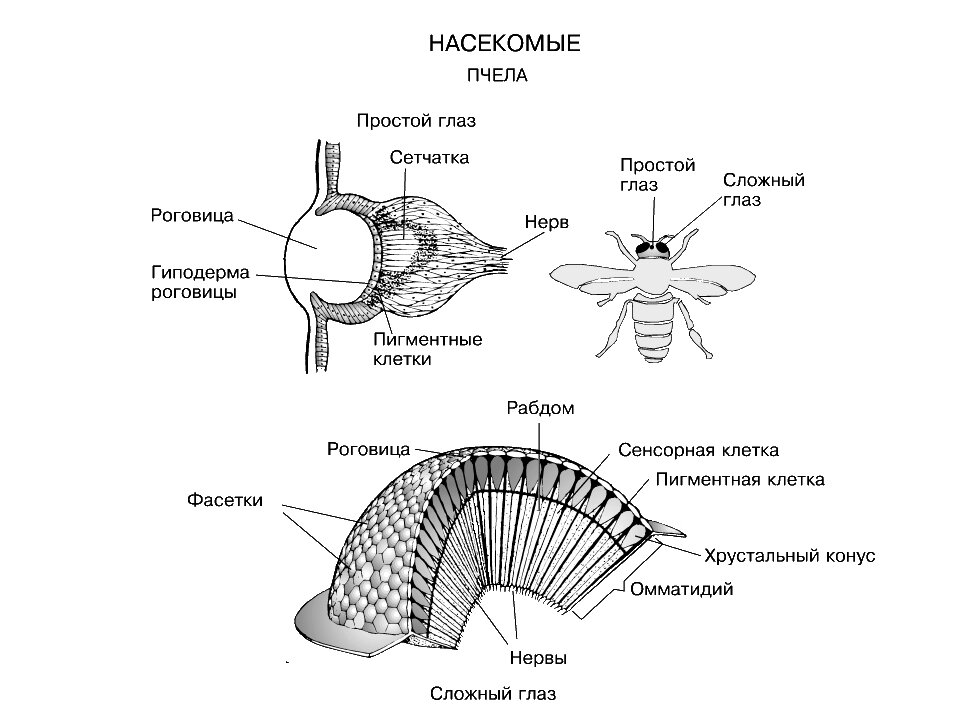 1. А. Онегов Русский мед / А. Онегов. - Москва: СПб. [и др.] : Питер, 1997. - 442 c. 2. А.М. Смирнов Болезни и вредители медоносных пчел / А.М. Смирнов. - М.: Книга по Требованию, 2012. - 334 c. 3. А.Н. Рыбальченко Загадка пчелиного роя / А.Н. Рыбальченко. - М.: Книга по Требованию, 2012. - 128 c. 4. В. Смирнов Ваш домашний помощник. Пчелы и пчеловодство / В. Смирнов. - М.: Рипол Классик, 2012. - 576 c. 5. В.Д. Лукоянов Пчеловодный инвентарь, пасечное оборудование / В.Д. Лукоянов, В.Н. Павленко. - М.: Агропромиздат, 1988. - 160 c. |