МЕТОДИЧКА-ЦИТОЛОГИЯ. Плазмолемма и другие клеточные мембраны Химические компоненты биомембран
 Скачать 239.81 Kb. Скачать 239.81 Kb.
|

|
Плазмолемма и другие клеточные мембраны Химические компоненты биомембран 1.Липидный компонент В основе любой биомембраны – липидный бислой: амфифильные липиды обращены в нём: гидрофобными частями – друг к другу, а гидрофильными – к водной фазе. 2. Белковый компонент а) Интегральные белки – насквозь пронизывают липидный бислой. б) Периферические белки – связаны с мембраной со стороны лишь одной из её поверхностей. 3. Углеводный компонент Углеводные компоненты обычно связаны - с липидами (в составе гликолипидов) - и с белками (в составе гликопротеинов) – в виде олигосахаридных цепей. Функции плазмолеммы Опорная функция: - изнутри к плазмолемме крепится цитоскелет, - снаружи плазмолемма взаимодействует с межклеточным веществом, что тоже может фиксировать клетку. Рецепторная функция: на внешней поверхности плазмолеммы могут находиться белки-рецепторы к гормонам, медиаторам и прочим сигнальным веществам. Взаимодействие с другими клетками – с помощью опять-таки рецепторов, а также адгезивных белков. Барьерная функция: за счёт липидного бислоя мембраны непроницаемы для гидрофильных и особенно заряженных соединений. Создание трансмембранного потенциала возбудимых клеток. Трансмембранный транспорт а) Способы помолекулярного (поионного) транспорта веществ В этом случае молекулы (или ионы) вещества проходят через мембрану относительно независимо друг от друга. Так могут транспортироваться только низкомолекулярные вещества.
б) Способы мультимолекулярного транспорта веществ Здесь за один акт переноса перемещается сразу большое число молекул. Так могут переноситься как низкомолекулярные, так и высокомолекулярные вещества.
Классификация органоидов Мембранные органоиды а) Двухмембранные органеллы 1. Ядра 2. Митохондрии б) Одномембранные органеллы или вакуолярная система клетки 1. Эндоплазматическая сеть (ЭПС): гранулярная (шероховатая) и гладкая 2. Комплекс (аппарат) Гольджи 3. Эндосомы и лизосомы 4. Пероксисомы 2. Безмембранные органеллы Гранулярные органеллы: рибосомы Фибриллярные органеллы: 1. Сократительные органеллы – миофибриллы, состоящие из миофиламентов. 2. Элементы цитоскелета: - микрофиламенты (МФ), - промежуточные филаменты, - микротрубочки (МТ) Производные элементов цитоскелета Производное микрофиламентов: 1.Каркас микроворсинок Производные микротрубочек 1. Центриоли, обычно расположенные парой (диплосомой). 2. Веретено деления. 3. Аксонема – каркас ресничек и жгутиков Одномембранные органоиды ЭПС представляет собой совокупность мембранных вакуолей, трубочек и плоских мешков (цистерн), распределённых в цитоплазме. Гладкая ЭПС. В гладкой ЭПС - цистерн почти нет, а с мембранами вакуолей и трубочек никакие видимые структуры не связаны. Функции: в мембранах гладкой ЭПС встроены ферменты гидроксилирования (или микросомального окисления), которые необходимы при синтезе многих липидов (например, стероидных гормонов), и для обезвреживания токсических соединений. Поэтому гладкая ЭПС особенно выражена в клетках надпочечников и гонад, синтезирующих стероидные гормоны, и печени в связи с синтезом холестерина и детоксикационной функцией гепатоцитов. Саркоплазматическая сеть – разновидность гладкой ЭПС. В скелетной и в сердечной мышечных тканях гладкая ЭПС обозначается как саркоплазматическая сеть. В ней хорошо развиты цистерны, в которых аккумулируются ионы Са2+ (с помощью Са2+-насоса). При возбуждении в цистернах открываются Са2+-каналы, и ионы Са2+ выходят в саркоплазму, стимулируя сокращение миофибрилл. Шероховатая (гранулярная) ЭПС. ЭПС представляет собой совокупность мембранных вакуолей, трубочек и плоских мешков (цистерн), распределённых тем или иным способом в цитоплазме. С мембранами данной сети со стороны гиалоплазмы связаны многочисленные рибосомы, что придаёт сети шероховатый вид. Функции: На этих (мембраносвязанных) рибосомах в соответствующих клетках образуются следующие белки: экспортные (выводящиеся из клетки) – белковые гормоны, пищеварительные ферменты, антитела и т.п., а также белки и ферменты мембран, лизосом и пероксисом самой клетки. Стадии синтеза белков: 1) трансляция мРНК на рибосомах, причём, синтезируемые пептидные цепи проникают через мембрану в полость ЭПС; 2) фолдинг (сворачивание) новообразованных белков; 3) начало модификации белков (связывание олигосахаридов и т.п.) Аппарат Гольджи. Аппарат Гольджи – постоянный партнёр шероховатой ЭПС в образовании всё тех же видов белков: экспортных, мембранных, лизосомных и пероксисомных. Состоит из диктиосом - скопление 5-10 плоских мембранных цистерн, лежащих параллельно друг другу. Диктиосомы соединены с шероховатой ЭПС и друг с другом трубочками и цистернами. Вся совокупность диктиосом и составляет комплекс Гольджи. Функции: В комплексе Гольджи происходят процессы, продолжающие те, что начинались в шероховатой ЭПС: 1) завершается модификация белков, 2) белки сортируются на экспортные, мембранные, лизосомальные и пероксисомные, 3) при этом белки концентрируются и «упаковываются» в соответствующих структурах – транспортных пузырьках, будущих лизосомах и т.д., 4) затем включаются механизмы транспорта этих структур в соответствующих направлениях. Лизосомы. Лизосомы – мембранные пузырьки, содержащие ферменты гидролиза биополимеров – лизосомные гидролазы. Они активны в сильно кислой среде, которая создаётся в лизосомах за счёт деятельности протонных насосов. Функция: внутриклеточное переваривание макромолекул. Типы лизосом: а) первичные – вновь образованные, с гомогенным содержимым, б) вторичные – включают перевариваемый материал, отчего их содержимое неоднородно, в) телолизосомы (остаточные, или резидуальные, тельца) – содержат непереваренные остатки белков и липидов, которые уплотняются и превращаются в пигмент старения липофусцин. Гидролазные пузырьки и эндосомы а) Теперь первичную лизосому называют просто гидролазным пузырьком. Эти пузырьки, как и мелкие протонные пузырьки, отщепляются от аппарата Гольджи. б) А сливающиеся пиноцитозные пузырьки, диффундирующие навстречу от плазмолеммы, образуют эндосому (первичную лизосому). в) Слияние гидролазных пузырьков с эндосомой даёт позднюю эндосому, которая при дальнейшем снижении рН превращается в лизосому (по старом у – вторичную лизосому). Пероксисомы. Пероксисомы, как и лизосомы, – мембранные пузырьки, содержащие большой набор ферментов. Только это ферменты уже не гидролитических, а окислительно-восстановительных реакций. Ферменты пероксисом: 1) оксидазы аминокислот и ряда других веществ; причём в результате этих реакций образуется пероксид водорода, Н2О2 – опасный окислитель; 2) ферменты пероксисом, устраняющие Н2О2 из среды, – каталаза и пероксидазы. Образование новых пероксисом - На мембраносвязанных и свободных рибосомах синтезируются ферменты пероксисом; - они проникают внутрь существующих пероксисом, - и от последних отпочковываются новые пероксисомы. Рибосома. Рибосома состоит из двух субъединиц – большой и малой. Каждая из них представляет собой свёрнутый рибонуклеопротеидный тяж. В неработающем состоянии субъединицы диссоциированы. Они объединяются вместе лишь при наличии мРНК и активированных аминокислот. Функция: Рибосомы – безмембранные органеллы, которые осуществляют трансляцию мРНК, т.е. включают в строящуюся пептидную цепь аминокислотные остатки в соответствии с последовательностью кодонов мРНК. Полисомы: Как правило, цепь мРНК одновременно транслируют несколько (до 5-6) рибосом, которые ползут друг за другом с определённым интервалом. Такие структуры называются полисомами (или полирибосомами). Типы рибосом: -цитоплазматические рибосомы - митохондриальные рибосомы, на которых образуется лишь малая часть белков митохондрий. Свободные и мембрано-связанные рибосомы А цитоплазматические рибосомы могут функционировать - в гиалоплазме (в свободном, мембранонесвязанном, состоянии) - и на мембранах ЭПС – на стороне, обращённой к гиалоплазме; это т.н. мембраносвязанные рибосомы. Очевидно, термин «мембраносвязанные рибосомы (или, точнее, полисомы)» означает ту же структуру, что и термин «шероховатая ЭПС». Следовательно, на этих рибосомах образуются те четыре группы белков, которые мы упоминали выше: экспортные, мембранные, лизосомальные и пероксисомные. Остальные белки (т.е. основная масса белков клетки) образуются свободными рибосомами (полисомами). Митохондрий. Митохондрии – двухмембранные структуры. Наружная мембрана митохондрий содержит широкие гидрофильные каналы и поэтому проницаема для многих низкомолекулярных веществ. Внутренняя мембрана имеет многочисленные впячивания – кристы, – которые значительно увеличивают её площадь. В эту мембрану встроены ферменты т.н. цепи переноса электронов (от окисляемых субстратов на кислород), и с переносом электронов по данной цепи сопряжено образование АТФ. Функции: - заключительные стадии окислительного распада питательных веществ, в т.ч. – цикл Кребса и β-окисление жирных кислот, - а также преобразование энергии окислительных реакций в энергию АТФ – т.н. окислительное фосфорилирование. Митохондрии, как полагают, произошли от древних аэробных бактерий, вступивших в симбиоз с эукариотическими клетками. Поэтому в них сохранилась система автономного синтеза белков, сходная с бактериальной: 1) собственная ДНК (мтДНК) – кольцевая, от 1 до 50 копий в органелле, включающая 37 генов; 2) собственные рибосомы, меньшие по размеру цитоплазматичских. Однако эта система обеспечивает синтез лишь 5% белков митохондрий. Цитоскелет и его производные а) Микрофиламенты (МФ) МФ образованы глобулярным белком актином: микрофиламент – двойная спираль (диаметром 5-7 нм), каждая нить которой представляет собой длинную цепочку из шарообразных молекул актина. Функции: а) В клетках МФ, ориентируясь, в основном, вдоль их длинной оси, образуют густую сеть. Особенно она выражена на периферии клеток, где препятствует их сильной деформации. б) Изменение длины МФ лежит в основе изменения формы клеток и различных видов клеточного движения. Участие по всасывании веществ МФ образует также цитоскелет микроворсинок – в частности, на апикальной поверхности эпителиальных клеток тонкой кишки. В центре микроворсинки – пучок из 40 МФ, а в основании микроворсинки МФ контактируют с молекулами миозина. Скольжение МФ относительно миозина способствует перемещению всасываемых веществ в тело клетки. б) Промежуточные филаменты - по диаметру (10 нм) находятся между МФ (5-7 нм) и МТ (24 нм), с чем и связано их название. Белковая природа данных филаментов зависит от вида ткани. В эпителии это кератин. в) Микротрубочки (МТ) Стенка МТ образована одним слоем глобулярного белка тубулина. На поперечном срезе – 13 глобул этого белка, образующих почти окружность (на самом деле укладка глобул в стенке – спиралевидная). Изменение длины Длина МТ (как и МФ) может меняться. Регулятором, видимо, служат специальные стабилизирующие их белки. При этом у МТ различают два конца: - минус-конец, часто закреплённый на одном из сателлитов (каковыми являются особые белковые тельца), - и плюс-конец, с которого и происходит рост (путём самосборки) или укорочение МФ. Функции: 1) МТ тоже образуют в клетке густую сеть, поддерживая форму клеток. 2) МТ выполняют и транспортную роль: например, в длинных отростках нервных клеток вдоль МФ, как по рельсам, перемещаются секреторные и иные гранулы. г) Производные МТ Центриоли. а) Центриоли образованы микротрубочками по формуле (9×3)+0. Это означает, что центриоль содержит 9 периферических триплетов МТ, расположенных по окружности, а в центре МТ нет. б) Центриоли располагаются парой – под прямым углом друг к другу. Такая структура называется диплосомой. в) В свою очередь, вокруг центриоли – т.н. центросфера, содержащая сателлиты, от которых отходят МТ цитоскелета. г) Вместе центриоли и центросфера составляют клеточный центр. Аксонема. а) Аксонема, или осевая нить, является цитоскелетом ресничек и жгутиков. Схема расположения в ней МТ такова: (9×2)+2. Т.е. имеется 9 периферических дуплетов МТ и одна центральная пара. б) Периферические дуплеты связаны друг с другом «ручками» из белка динеина, а с центральной парой – радиальными мостиками. в) При замыкании и размыкании динеиновых мостиков соседние дуплеты скользят относительно друг друга, что приводит к изгибу (биению) аксонемы. 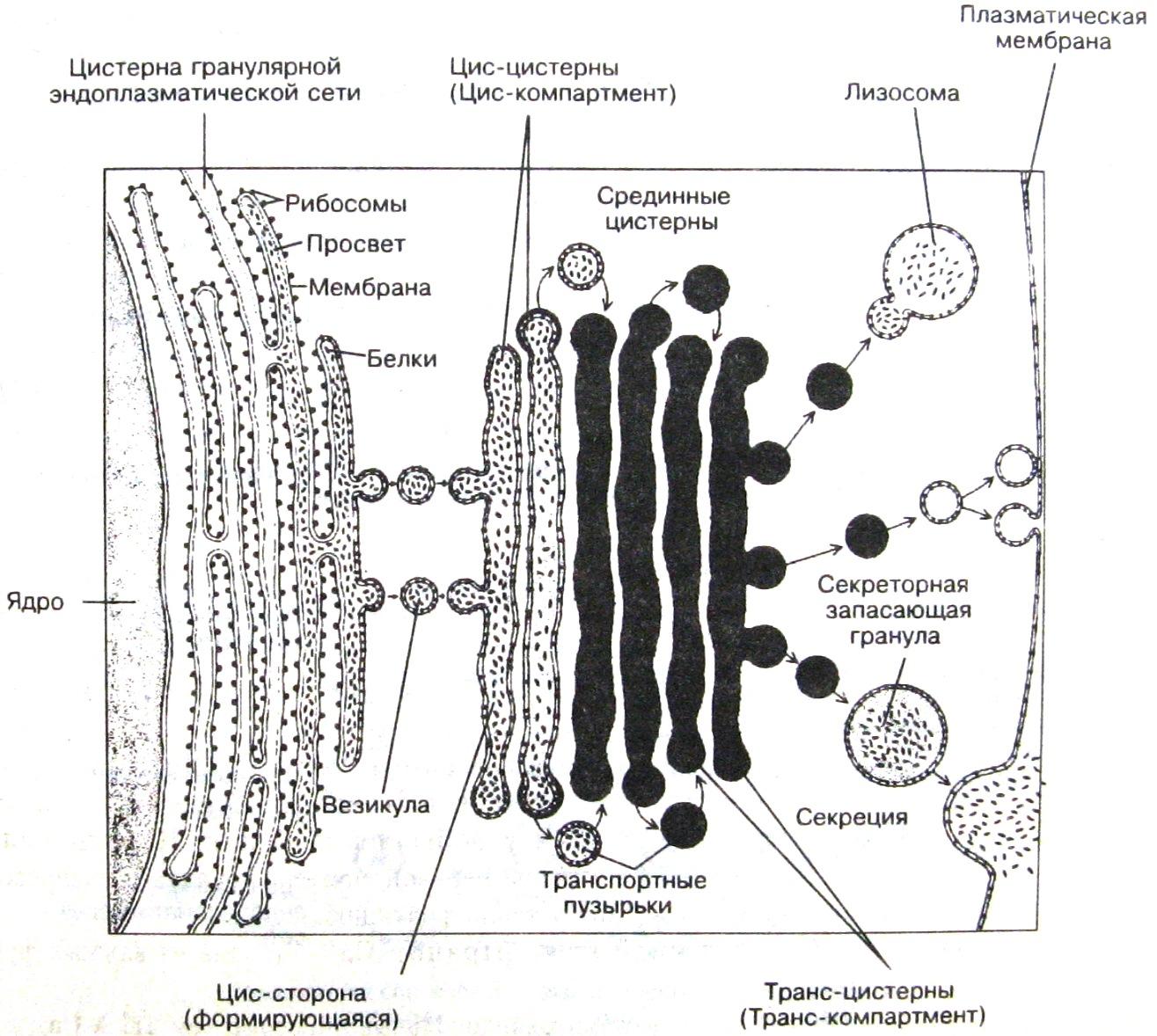 Компоненты ядра клетки: - ядерная оболочка, - внешне бесструктурный ядерный матрикс, - наследственный материал, представленный хромосомами и воспринимающийся в интерфазе как хроматин, а в митозе – как кариотип, - и ядрышки. Ядерная оболочка Ядерная оболочка состоит из двух мембран, разделённых перинуклеарным пространством и образующих полый двухслойный мешок. Наружная мембрана – это компонент цитоплазматической ЭПС, с которым могут быть связаны рибосомы. Внутренняя мембрана связана со структурами самого ядра: а именно – с прилегающей к ней ядерной пластинкой (ламиной), к которой крепятся концы всех хромосом, причём, в строго определённых местах. Ядерные поры. Ядерная оболочка пронизана большим количеством (порядка 2000-4000) крупных пор. В области краёв поры наружная и внутренняя мембраны сливаются, образуя округлое отверстие диаметром 120 нм. В это отверстие встроен т.н. комплекс поры, в котором имеются каналы уже гораздо меньшего, но всё же значительного, диаметра – 10 нм. Транспорт через поры а) Через каналы происходит интенсивный транспорт молекул и крупных частиц в обоих направлениях: - из цитоплазмы в ядро – нуклеотидов, хромосомных и рибосомных белков, регуляторных факторов; - и из ядра в цитоплазму – многочисленных мРНК, тРНК, рибосомных субъединиц. б) Чаще всего используется, видимо, активный транспорт – в случае крупных молекул и частиц. В этом участвуют специальные рецепторные белки и используется энергия АТФ. Ядерный матрикс Несмотря на внешнюю бесструктурность, матрикс ядра включает не только жидкую часть – ядерный сок (кариоплазму), но и белковый каркас (кариоскелет): * упоминавшуюся ядерную пластинку (ламину), через которую концы хромосом крепятся к внутренней ядерной мембране, * а также внутриядерную фибриллярную сеть – для фиксации хромосом и разнообразных белковых комплексов. 3. Хромосомы и хроматин 1) ДНК, хроматиды и хромосомы ДНК. В ядре любой диплоидной по ДНК клетки человека – 46 молекул ДНК. Средняя длина одной из них – 4 см, общая протяжённость – 185 см. Хроматиды. Каждая молекула ДНК связана с определёнными белками и образует дезоксирибонуклеопротеидный тяж – хроматиду. Хромосома. а) Большую часть времени существования клетки каждая хроматида представляет собой отдельную хромосому. б) Перед делением клетки - происходит удвоение ДНК и, соответственно, хроматид; - число хромосом остаётся прежним, но каждая из них теперь содержит по 2 хроматиды и, соответственно, по 2 мол. ДНК; - всего в ядре клетки оказывается 92 молекулы ДНК. 2) Интерфазные хромосомы и хроматин Интерфазные хромосомы В неделящейся клетке, а также в клетке, которая только готовится к делению, но ещё не делится (в т.ч. на стадии удвоения ДНК), хромосомы не находятся в степени максимальной конденсации и поэтому при световой микроскопии неразличимы. Но при этом они сохраняют свою индивидуальность и определенную упорядоченность расположения: например, крепятся концами к строго определённым участкам внутренней ядерной мембраны. Такие хромосомы называются интерфазными. Совокупность интерфазных хромосом обозначается как хроматин. Выделяют два типа хроматина. а) Гетерохроматин – это относительно (хотя и не полностью) конденсированные, а потому функционально неактивные, хромосомы и (или) части хромосом. Под световым микроскопом воспринимаются как тёмные глыбки. б) Эухроматин – это относительно (хотя и не полностью!) деконденсированные, т.е. функционально активные, хромосомы и (или) части хромосом. На световом уровне остаются практически неокрашенными. Виды гетерохроматина В свою очередь, гетерохроматин подразделяют на а) факультативный – способный превращаться в эухроматин; б) конститутивный – никогда и ни в одной клетке к подобному превращению не способный. Пример конститутивного гетерохроматина – половой хроматин, или тельце Бара, – та из двух Х-хромосом у женщин, которая всегда находится в конденсированном состоянии. 3. Мета- и анафазные хромосомы и кариотип Полной конденсации хромосомы достигают лишь в процессе деления – к началу метафазы. При этом они приобретают форму, строго определённую для каждой пары гомологичных хромосом. а) Метафазные хромосомы состоят из двух хроматид, которые начали расходиться, но ещё связаны в центромерной области. б) Анафазные хромосомы – это уже разошедшиеся хроматиды. Совокупность числа, размеров и особенностей строения метафазных (или анафазных) хромосом обозначается как кариотип. Таким образом, отличия от хроматина состоят, - во-первых, в том, что последний (хроматин) образован интерфазными хромосомами, а кариотип – мета- или анафазными, - а во-вторых, в том, что в кариотипе хромосомы характеризуются формой и размерами. Отделы хромосом Части, выявляющиеся в мета- и анафазных хромосомах: а) центромера (первичная перетяжка), в этой области находится кинетохор – место прикрепления веретена деления; б) плечи – части хромосомы по сторонам от центромеры; в) теломеры – конечные участки плеч. У некоторых хромосом в одном из плеч имеется и вторичная перетяжка. Это место расположения генов рибосомных РНК. По положению центромеры хромосомы делят на три вида: а) метацентрические – с равными плечами, б) субметацентрические – с плечами неодинаковой длины, в) акроцентрические – одно плечо практически отсутствует. Белки хромосом В структурных преобразованиях хромосом, а также в процессах их функционирования очень важную роль играют белки. Хромосомные белки подразделяют на оснóвные и кислые. Оснóвные белки представлены гистонами пяти видов. а) Оснóвность им придаёт высокое содержание лизина и аргинина, чьи радикалы имеют положительный заряд. Это позволяет им взаимодействовать с отрицательно заряженными фосфатными группами ДНК. б) Также в гистонах много аминокислот с гидрофобными радикалами. Благодаря этому, гистоны хорошо взаимодействуют и друг с другом. в) Именно гистоны в первую очередь отвечают за структурное состояние хромосом. На кислые (негистоновые) белки приходится только 20-40% массы хромосомных белков (остальное – гистоны). Тем не менее, в отличие от гистонов, они чрезвычайно разнообразны: Их функции: - одни кислые белки участвуют в образовании высших уровней укладки хромосом (структурная функция), - другие являются ферментами тех важнейших процессов, в которых участвуют хромосомы; - третьи – регулируют активность ферментов и доступность различных участков ДНК. Уровни организации хромосом а) Количество уровней укладки при разных состояниях хромосом Различают 4 уровня компактизации хромосом. а) Эухроматин обладает только одним, самым низшим уровнем (нуклеосомным), что сохраняет его матричную активность. б) Гетерохроматин имеет уже 3 уровня упаковки (от нуклеосомного до хромомерного), что лишает его активности, но всё ещё не позволяет различить отдельные хромосомы. в)А в метафазных (и анафазных) хромосомах - другой, более компактный, третий уровень организации (хромонемный вместо хромомерного) - и, кроме того, добавляется ещё один – четвёртый уровень (хроматидный). Так что, в итоге, длина ДНК (и хромосомы) сокращается в 10.000 раз.
б) Характеристика уровней компактизации хромосом
Ядрышки Ядрышки – последние из перечисленных в начале темы компонентов ядра. Ядрышко (нуклеола) – самая плотная структура ядра, обычно имеющая округлую форму. Нередко в ядре – не одно, а несколько ядрышек. При этом ядрышки – не самостоятельные структуры: они являются производными хроматина. Всего в ядрышке различают три компонента: 1) ядрышковый организатор – совокупность некоего числа копий генов рибосомальных РНК, 2) фибриллярный компонент – синтезируемые на этих генах пре-рРНК и созревающие из цепи рРНК, 3) а также глобулярный компонент – формирующиеся здесь же субъединицы рибосом. Ядрышковый организатор а) Гены рибосомальных РНК локализуются в пяти парах хромосом, т.е. в 10 хромосомах (в области их вторичной перетяжки). б) При этом гены трёх видов рРНК (из четырёх) расположены друг за другом («цугом»), образуя т.н. кластер, который считывается при транскрипции как единое целое. в) В каждом ядрышковом организаторе – несколько сотен или даже тысяч копий как этого кластера, так и не входящего в него гена четвёртого вида (5 S-) рРНК. г) Количество ядрышек в ядре зависит от того, во сколько групп объединились 10 ядрышковых организаторов – в одну или несколько Фибриллярный и глобулярный компоненты а) Превращение пре-рРНК в зрелые рРНК сводится к разрезанию её на отдельные рРНК и к дополнительной модификации последних. б) Зрелые рРНК вовлекаются тут же – в ядрышках – в процесс самосборки субъединиц рибосом, в котором участвует и несколько десятков рибосомных белков, поступающих из цитоплазмы. Функции ядер и внутриядерные процессы Ядра клеток, в целом, выполняют три основные функции: 1) обеспечивают использование информации ДНК в самих клетках, 2) сохраняют наследственный материал для его продолжительного функционирования и для передачи его дочерним клеткам, 3) в определённых условиях, напротив, уничтожают свой геном. В рамках реализации первой функции в ядрах проходят процессы: а) транскрипция определённых участков ДНК (синтез пре-мРНК), б) созревание (процессинг) мРНК, в) синтез и созревание тРНК и рРНК, сборка субъединиц рибосом, г) сложная регуляция активности разных участков ДНК. Вторую функцию ядер обеспечивают процессы: а) репарация ДНК (обнаружение дефектов ДНК и их исправление), б) репликация (удвоение) ДНК и других компонентов хромосом, в) конденсация хромосом перед делением. Третья функция ядер реализуется в ходе апоптоза – клеточного «самоубийства». Деления клеток а) Способность клеток к делениям: по этому признаку различают 3 группы клеток.
б) Митотический цикл: в нём выделяют 4 периода интерфаза: G1-период – постмитотический, или пресинтетический Это интервал времени от образования клетки (путём митоза) до начала синтеза в ней ДНК (и хромосомных белков). Происходит восстановление содержания белков цитоплазмы и рост клетки. В ядре – 46 хромосом по 1-й хроматиде в каждой; значит, и 46 молекул ДНК. Именно в G1-периоде определяется, войдёт ли клетка в митотический цикл или нет. Однако ближе к концу периода наступает момент (точка рестрикции», после которого выбор уже не происходит, то есть клетка всё равно войдёт в S-период, даже если на неё перестанут действовать митогенные факторы. S-период – синтетический В ядре происходит репликация ДНК и дупликация центриолей. Количество ДНК постепенно возрастает: от 46 до 92. G2-период – премитотический, или постсинтетический В этот период синтезируются вещества, необходимые для митоза: в т.ч. белок микротрубочек тубулин для формирования веретена деления. В ядре – 46 хромосомы по 2 хроматиды в каждой, то есть 92 молекулы ДНК. Митоз Это центральное событие цикла, в ходе которого тетраплоидная по ДНК клетка делится на 2 диплоидные клетки. Примерная продолжительность периодов для быстро делящихся клеток такова: весь цикл – 24 часа, в т.ч G1-период – 9 ч, S-период – 10 ч, G2-период – 4,5 ч, M – 0,5 ч. Профаза а) Конденсация хромосом (формирование четырёх уровней укладки). б) Вследствие этого – исчезновение ядрышек и прекращение синтеза РНК. в) Разрушение ядерной ламины; распад мембран ядра, ЭПС и аппарата Гольджи на мелкие мембранные пузырьки. г) Расхождение центриолей (диплосом) к полюсам клетки и начало формирования веретена деления. Метафаза а) Хромосомы достигают максимальной конденсации, а также образуют в экваториальной плоскости т.н. метафазную звезду и почти по всей длине (кроме центромер) разделяются на хроматиды. б) Веретено деления включает микротрубочки (МТ), идущие от каждой из двух диплосом в трёх направлениях: - кинетохорные МТ – к кинетохоре одной из хроматид каждой хромосомы, - полярные МТ – к экватору клетки, где контактируют с таки же МТ от противоположного полюса; - астральные – к близлежащим областям плазмолеммы. в) MPF фосфорилирует и тем самым активирует ещё один белок – анафазу обеспечивающий фактор (АPС – anaphase-promotingcomplex). Анафаза Хромосомы окончательно разделяются на хроматиды, и последние расходятся по разным полюсам клетки. Этому способствуют изменение длины МТ – укорочение кинетохорных и удлинение полярных МТ, а также белки-транслоказы, перемещающие хроматиды вдоль кинетохорных МТ. Телофаза а) Большинство событий телофазы – это эффекты «отмены» MPF. Поэтому они, в основном, обратны событиям профазы: - из мембранных пузырьков образуются ядерные мембраны двух дочерних ядер, а также мембраны ЭПС и аппарата Гольджи; - хромосомы деконденсируются, и вновь формируются ядрышки. б) Кроме того, происходит цитотомия (цитокинез): актомиозиновое кольцо сжимается между двумя половинами делящейся клетки (образуя перетяжку) и, в конечном счёте, разделяет клетку на две дочерние. |