По своей химической структуре гормоны можно разделить на 3 группы
 Скачать 320.33 Kb. Скачать 320.33 Kb.
|

|
Гормо́ны — биологически активные вещества органической природы, вырабатывающиеся в специализированных клетках желёз внутренней секреции, поступающие в кровь, связывающиеся с рецепторами клеток-мишеней и оказывающие регулирующее влияние на обмен веществ и физиологические функции. Гормоны служат гуморальными (переносимыми с кровью) регуляторами определённых процессов в различных органах. По своей химической структуре гормоны можно разделить на 3 группы: 1) белки и пептиды (например, гормоны поджелудочной железы – инсулин и глюкагон; гормоны гипофиза и гипоталамуса); 2) производные аминокислот (например, гормоны мозгового вещества надпочечников – адреналин и норадреналин); 3) стероидные гормоны, т.е. производные липидов (например, большинство половых гормонов, глюкокортикостероиды). Физиологическое действие гормонов направлено на: 1) обеспечение гуморальной, т.е. осуществляемой через кровь, регуляции биологических процессов; 2) поддержание целостности и постоянства внутренней среды, гармоничного взаимодействия между клеточными компонентами тела; 3) регуляцию процессов роста, созревания и репродукции. Гормоны регулируют активность всех клеток организма. Они влияют на остроту мышления и физическую подвижность, телосложение и рост, определяют рост волос, тональность голоса, половое влечение и поведение. Благодаря эндокринной системе человек может приспосабливаться к сильным температурным колебаниям, излишку или недостатку пищи, к физическим и эмоциональным стрессам. Изучение физиологического действия эндокринных желез позволило раскрыть секреты половой функции и чудо рождения детей, а также ответить на вопрос, почему одни люди высокого роста, а другие низкого, одни полные, другие худые, одни медлительные, другие проворные, одни сильные, другие слабые. Действие гормонов на клеточном уровне осуществляется по двум основным механизмам: не проникающие в клетку гормоны (обычно водорастворимые) действуют через рецепторы на клеточной мембране, а легко проходящие через мембрану гормоны (жирорастворимые) – через рецепторы в цитоплазме клетки. Во всех случаях только наличие специфического белка-рецептора определяет чувствительность клетки к данному гормону, т.е. делает ее «мишенью». Первый механизм действия, подробно изученный на примере адреналина, заключается в том, что гормон связывается со своими специфическими рецепторами на поверхности клетки; связывание запускает серию реакций, в результате которых образуются т.н. вторые посредники, оказывающие прямое влияние на клеточный метаболизм. Такими посредниками служат обычно циклический аденозиномонофосфат (цАМФ) и/или ионы кальция; последние высвобождаются из внутриклеточных структур или поступают в клетку извне. И цАМФ, и ионы кальция используются для передачи внешнего сигнала внутрь клеток у самых разнообразных организмов на всех ступенях эволюционной лестницы. Однако некоторые мембранные рецепторы, в частности рецепторы инсулина, действуют более коротким путем: они пронизывают мембрану насквозь, и когда часть их молекулы связывает гормон на поверхности клетки, другая часть начинает функционировать как активный фермент на стороне, обращенной внутрь клетки; это и обеспечивает проявление гормонального эффекта. Второй механизм действия – через цитоплазматические рецепторы – свойствен стероидным гормонам (гормонам коры надпочечников и половым), а также гормонам щитовидной железы (T3 и T4). Проникнув в клетку, содержащую соответствующий рецептор, гормон образует с ним гормон-рецепторный комплекс. Этот комплекс подвергается активации (с помощью АТФ), после чего проникает в клеточное ядро, где гормон оказывает прямое влияние на экспрессию определенных генов, стимулируя синтез специфических РНК и белков. Именно эти новообразованные белки, обычно короткоживущие, ответственны за те изменения, которые составляют физиологический эффект гормона. БИОСИНТЕЗ ТРИГЛИЦЕРИДОВ Известно, что скорость биосинтеза жирных кислот во многом определяется скоростью образования триглицеридов и фосфолипидов, так как свободные жирные кислоты присутствуют в тканях и плазме крови в небольших количествах и в норме не накапливаются. Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтеза триглицеридов в тканях протекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения. В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата: Глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с процессами гликолиза и гликогенолиза. Известно, что в процессе гли-колитического распада глюкозы образуется дигидроксиацетонфосфат (см. главу 10). Последний в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат: Дигидроксиацетонфосфат в присутствии цитоплазматической глицерол-3-фосфатдегидрогеназы способен превращаться в глицерол-3-фосфат Отмечено, что если содержание глюкозы в жировой ткани понижено (например, при голодании), то образуется лишь незначительное количество глицерол-3-фосфата и освободившиеся в ходе липолиза свободные жирные кислоты не могут быть использованы для ресинтеза триглицеридов, поэтому жирные кислоты покидают жировую ткань. Напротив, активация гликолиза в жировой ткани способствует накоплению в ней триглицеридов, а также входящих в их состав жирных кислот. В печени наблюдаются оба пути образования глицерол-3-фосфата. Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты (т.е. «активными» формами жирной кислоты – ацил-КоА). В результате образуется фосфатидная кислота (фосфатидат): Образование фосфатидной кислоты из глицерол-3-фосфата Как отмечалось, ацилирование глицерол-3-фосфата протекает последовательно, т.е. в 2 этапа. Сначала глицерол-3-фосфат-ацилтрансфераза катализирует образование лизофосфатидата (1-ацилглицерол-3-фосфата, а затем 1-ацилглицерол-3-фосфат-ацилтрансфераза катализирует образование фосфатидата (1,2-диацилглицерол-3-фосфата) . Далее фосфатидная кислота гидролизуется фосфатидат-фосфогидро-лазой до 1,2-диглицерида (1,2-диацилглицерола): Образование 1,2-диглицерида из фосфатидной кислоты Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол). Эта реакция катализируется диацилглицерол-ацилтрансферазой: Образование триглицерида из 1,2-диглицерида Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов в стенке тонкой кишки из β-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления пищевых жиров. На рис. 11.6 представлены глицерофосфатный, дигидроксиацетон-фосфатный и β-моноглицеридный (моноацилглицероловый) пути синтеза триглицеридов. 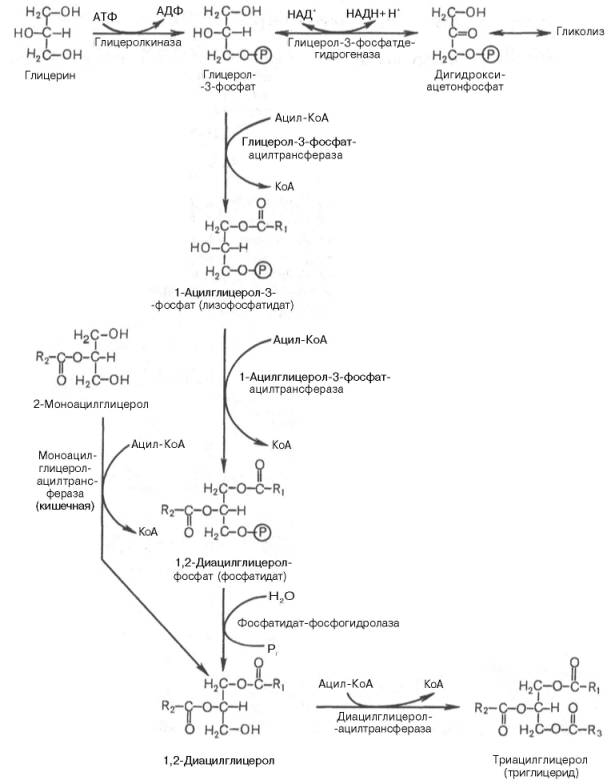 Рис. 11.6. Биосинтез триглицеридов (триацилглицеролов). Установлено, что большинство ферментов, участвующих в биосинтезе триглицеридов, находятся в эндоплазматическом ретикулуме, и только некоторые, например глицерол-3-фосфат-ацилтрансфераза,– в митохондриях. В отличие от триглицеридов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды играют важную роль в структуре и функции клеточных мембран, активации мембранных и лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в цепи «дыхательных» ферментов. Особая роль фосфолипидам отводится в формировании липо-протеидных комплексов. Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазма-тической сети клетки. Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфин-гомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов. Биосинтез фосфатидилэтаноламина. Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфоэтаноламина: Биосинтез фосфатидилэтаноламина. Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфоэтаноламина 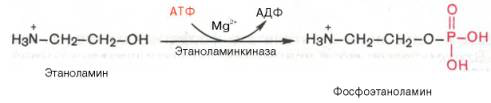 Затем фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (PPi): Фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (PPi) 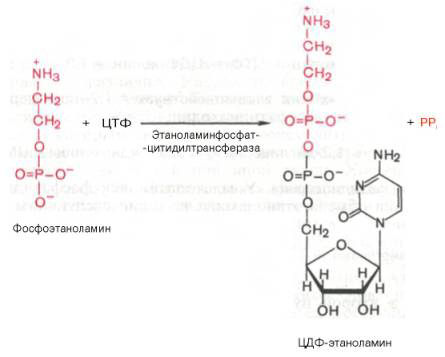 В следующей реакции ЦДФ-этаноламин, взаимодействуя с 1,2-дигли-церидом, образующимся при дефосфорилировании фосфатидной кислоты, превращается в фосфатидилэтаноламин. Реакция катализируется ферментом этаноламинфосфотрансферазой: ЦДФ-этаноламин + 1,2-диглицерид –> Фосфатидилэтаноламин + ЦМФ. Биосинтез фосфатидилхолина (лецитина). Фосфатидилэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метальных групп от трех молекул S-аденозилметионина (донор метальных групп, см. главу 6) к аминогруппе остатка этаноламина образуется фосфатидилхолин: Образование фосфатидилхолина из фосфатидилэтаноламина Существует еще один путь синтеза фосфатидилхолина в клетках животных. В этом случае, как и при синтезе фосфатидилэтаноламина, используется ЦТФ в качестве переносчика, но уже не фосфоэтаноламина, а фос-фохолина. На первом этапе синтеза свободный холин активируется под действием холинкиназы с образованием фосфохолина: 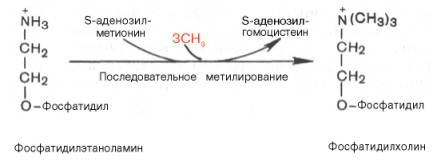 Холин + АТФ –> Фосфохолин + АДФ. Затем фосфохолин реагирует с ЦТФ, образуя цитидиндифосфатхолин (ЦДФ-холин): Фосфохолин + ЦТФ –> ЦДФ-холин + РРi. В дальнейшем ЦДФ-холин взаимодействует с 1,2-диглицеридом, в результате чего образуется фосфатидилхолин: ЦДФ-холин + 1,2-диглицерид –> Фосфатидилхолин + ЦМФ. Биосинтез фосфатидилсерина. У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем: Существует и второй путь образования фосфатидилсерина, который связан с предварительным вовлечением фосфатидной кислоты в синтез фосфоглицеридов: 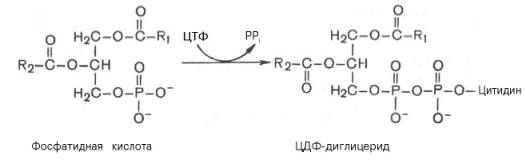 ЦДФ-диглицерид из фосфатидной кислоты Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина: ЦДФ-диглицерид + L-серин –> Фосфатидилсерин + ЦМФ. Таким же путем образуется фосфатидилинозитол. Биосинтез сфингомиелина. Интермедиатом в биосинтезе сфингомиелина является церамид (N-ацилсфингозин), который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия (реакции) церамида с ЦДФ-холином: Интермедиатом в биосинтезе сфингомиелина является церамид (N-ацилсфингозин), который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия (реакции) церамида с ЦДФ-холином Следует отметить, что различие в синтезе холин- и этаноламинсодер-жащих фосфолипидов, с одной стороны, и инозитсодержащих фосфоли-пидов – с другой, заключается в том, что в первом случае при участии ЦТФ образуется ЦДФ-холин или ЦДФ-этаноламин – реакционноспособные азотистые основания, а во втором случае при участии ЦТФ образуется ЦДФ-диглицерид – реакционноспособная форма диглицерида. Белки при взаимодействии с некоторыми химическими веществами дают окрашенные соединения. Образование этих соединений происходит при участии радикалов аминокислот, их специфических групп или пептидных связей. Цветные реакции позволяют установить наличие белка в биологическом объекте или растворе и доказать присутствие определенных аминокислот в белковой молекуле. На основе цветных реакций разработаны некоторые методы количественного определения белков и аминокислот. Универсальными считают биуретовую и нингидриновую реакции, так как их дают все белки. Ксантопротеиновая реакция, реакция Фоля и др. являются специфическими, так как они обусловлены радикальными группами определенных аминокислот в молекуле белка. БИУРЕТОВАЯ РЕАКЦИЯ. Реакция обусловлена наличием в белках, пептидах, полипептидах пептидных связей, которые в щелочной среде образуют с ионами меди (II) комплексные соединения, окрашенные в фиолетовый (с красным или с синим оттенком) цвет. Окраска обусловлена наличием в молекуле не менее двух групп -CO-NH-, связанных непосредственно между собой или при участии атома углерода или азота. РЕАКЦИЯ ФОЛЯ Это реакция на цистеин и цистин. При щелочном гидролизе «слабосвязанная сера» в цистеине и цистине достаточно легко отщепляется, в результате чего образуется сероводород, который, реагируя со щелочью, дает сульфиды натрия или калия. При добавлении ацетата свинца(II) образуется осадок сульфида свинца(II) серо-черного цвета. Описание опыта. В пробирку наливают 1 мл раствора цистина, прибавляют 0,5 мл 20%-го раствора гидроксида натрия. Смесь нагревают до кипения, а затем добавляют 0,5 мл раствора ацетата свинца(II). Наблюдается выпадение серо-черного осадка сульфида свинца(II): КСАНТОПРОТЕИНОВАЯ РЕАКЦИЯ Эта реакция используется для обнаружения a-аминокислот, содержащих ароматические радикалы. Тирозин, триптофан, фенилаланин при взаимодействии с концентрированной азотной кислотой образуют нитропроизводные, имеющие желтую окраску. В щелочной среде нитропроизводные этих a-аминокислот дают соли, окрашенные в оранжевый цвет. Описание опыта. В пробирку наливают 1 мл раствора тирозина и добавляют 0,5 мл концентрированной азотной кислоты. Смесь нагревают до появления желтой окраски. После охлаждения добавляют 1–2 мл 20%-го раствора гидроксида натрия до появления оранжевой окраски раствора: КСАНТОПРОТЕИНОВАЯ РЕАКЦИЯ Эта реакция используется для обнаружения a-аминокислот, содержащих ароматические радикалы. Тирозин, триптофан, фенилаланин при взаимодействии с концентрированной азотной кислотой образуют нитропроизводные, имеющие желтую окраску. В щелочной среде нитропроизводные этих a-аминокислот дают соли, окрашенные в оранжевый цвет. Описание опыта. В пробирку наливают 1 мл раствора тирозина и добавляют 0,5 мл концентрированной азотной кислоты. Смесь нагревают до появления желтой окраски. После охлаждения добавляют 1–2 мл 20%-го раствора гидроксида натрия до появления оранжевой окраски раствора: Реакция на триптофан(Адамкевича) Окраска возникает за счет реакции триптофана с глиоксиловой кислотой, всегда присутствующей в уксусной кислоте. Небольшое количество меди повышает чувствительность этой реакции. Реакция основана на способности триптофана в кислой среде реагировать с глиоксиловой кислотой с образованием соединения, окрашенного в красно- фиолетовый цвет. Для проведения реакции используют ледяную (концентрированную) уксусную кислоту, в которой всегда в небольшом количестве присутствует глиоксиловая кислота. В качестве водоотнимающего средства в реакции используется концентрированная серная кислота. |