Карташев Наумов Практикум по зоологии. "Практикум по зоологии позвоночных". Подтип бесчерепные acrania тема строение ланцетника систематическое положение объекта
 Скачать 5.56 Mb. Скачать 5.56 Mb.
|

|
5). Резко редуцируется скелет пальцев. Сохраняются две фаланги II пальца (рис. 111, 6), продолжающие ось пряжки. От I пальца сохраняется одна фаланга (рис. 111, 7), к ней прикрепляется пучок перьев, образующий крылышко (alula; см. рис. 98, 4). III палец представлен также одной фалангой (рис. 111, 8), прикрепленной к основанию первой фаланги II пальца. Эти преобразования в скелете кисти (образование carpometacarpus, редукция пальцев) обеспечивают прочную опору для первостепенных маховых перьев — части крыла, несущей в полете наибольшую нагрузку. Тазовый пояс состоит из трех пар срастающихся вместе костей: Широкая и длинная подвздошная кость (ilium; см. рис. 106, 15; рис. 107, 4) на большом протяжении срастается со ложным крестцом. К ее наружному краю прирастает седалищная кость (ischium; см. рис. 106, 16; рис. 107, 5), а ниже расположена направленная назад палочкообразная лобковая кость (pubis; см. рис. 106, 17; рис. 107, 6). Все три кости таза образуют вертлужную впадину (acetabulum; см. рис. 106, 18), в которую входит головка бедра. Большая поверхность тазовых костей и их прочное (неподвижное) соединение с осевым скелетом обеспечивают прочную опору для прикрепления мощных мышц. Эти особенности тазового пояса определяются тем, что передние конечности выполняют только функцию полета, а при движении по земле вся тяжесть тела поддерживается лишь задними конечностями. Лобковые и седалищные кости правой и левой сторон не соединяются друг с другом на брюшной поверхности (см. рис. 107). Поэтому таз птиц называют открытым; он дает возможность откладывать крупные яйца. Можно предполагать, что открытый таз, увеличивающий подвижность брюшной стенки в тазовой области, способствует интенсификации дыхания в полете. 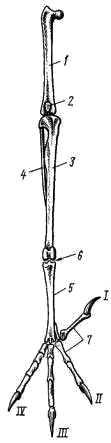 Рис. 112. Скелет задней конечности вороны: 1 — бедренная кость, 2 — коленная чашечка, 3 — голено-предплюсна (tibio-tarsus), 4 — малая берцовая кость, 5 — цевка (слившиеся кости предплюсны и плюсны), 6 – интертарзальный сустав, 7 — фаланги пальцев; I — IV — пальцы Скелет задней конечности представлен мощными трубчатыми костями. Бедро (femur; рис. 112, 1) на проксимальном конце имеет хорошо развитую округлую головку для сочленения с тазовым поясом. На дистальном конце образуются рельефные суставные поверхности для сочленения с костями голени. В области коленного сустава в мышечном сухожилии лежит коленная чашечка (patella; рис. 112, 2). Основной элемент голени — массивная большая берцовая кость, с нижним дистальным концом которой сливаются две проксимальные кости предплюсны; образуется костный комплекс, представляющий собой голено-предплюсну, или тибио-тарзус (tibio-tarsus; рис. 112, 3). Малая берцовая кость (fibula; рис. 112, 4) сильно редуцирована и в виде маленькой тоненькой косточки прирастает к верхней части наружной поверхности большой берцовой кости. Дистальные элементы предплюсны (tarsus) и все элементы плюсны (metatarsus) сливаются у птиц в единую кость — цевку, или плюсну-предплюсну (tarso-metatarsus; рис. 112, 5). Благодаря этому в конечности появляется добавочный рычаг. Подвижное сочленение располагается между двумя рядами костей предплюсны (между костями приросшими к голени, и костями, слившимися с плюсной), поэтому, как и у пресмыкающихся, этот сустав называется интертарзальным (рис. 112, 6). На дистальном конце цевки образуются хорошо выраженные суставные поверхности для причленения фаланг пальцев (phalanges digitorum; рис. 112, 7). У подавляющего большинства птиц в задней конечности развиваются четыре пальца, из которых I направлен назад, а II, III, IV — вперед. Мощные длинные кости скелета задних конечностей, появление добавочного рычага (образование tarso-metatarsus), резкий рельеф суставных поверхностей — все это обеспечивает прочность и подвижность задних конечностей. ЗАКЛЮЧЕНИЕ Морфологически класс птиц во многих чертах сохраняет большое сходство со своими предками — пресмыкающимися. На это указывает целый ряд признаков: почти полное отсутствие кожных желез, то, что дистальные отделы задних конечностей покрыты роговыми чешуями, роговой покров клюва, диапсидного типа череп (с редуцированной, однако, верхней дугой), имеющий только один затылочный мыщелок, состав костей черепа, то, что имеются шейные ребра и крючковидные отростки грудных ребер, открытый таз (как у некоторых ископаемых динозавров), большое сходство строения мочеполовой системы и периферических отделов кровеносной системы, присутствие клоаки, размножение откладыванием яиц, сходство эмбрионального развития и т. п. В процессе эволюции птицы обособились от рептилиеобразных предков путем многих, но часто относительно небольших преобразований морфологических структур, обеспечиваюших существенную интенсификацию физиологических функций и тем самым резко повысивших общий уровень организации птиц по сравнению с пресмыкающимися. Полет и высокая общая подвижность птиц связаны с большим расходом энергии, требующим быстрой компенсации. Поэтому в эволюции птиц большое значение имело становление особенностей организации, связанных не только с обеспечением физической возможности полета, но и с общим повышением интенсивности обмена веществ. Возможность полета птиц обеспечивается целым рядом прямых приспособлений. К ним относятся: компактность тела, жесткость и неподвижность туловищного отдела осевого скелета и редукция его хвостового отдела, резкое увеличение размеров грудины и образование на ней киля, прочность пояса передних конечностей (в том числе и образование специального амортизирующего устройства — вилочки), строение скелета крыла (включая и особенности суставных поверхностей и особенно резкие изменения в строении кисти), мощность мускулатуры, обеспечивающая движения крыла, развитие перьев — легких и прочных роговых образований, создающих несущие плоскости крыльев и хвоста и обеспечивающих обтекаемость тела в по лете, редукция зубов, облегчающая скелет черепа, общее увеличение объема головного мозга и особенно сильное развитие мозжечка — центра равновесия и координации движений, увеличение массы нервных клеток спинного мозга в области крыльев и т. д. Клюв с его подвижным скелетным механизмом и дифференцированной мускулатурой стал очень тонко действующим инструментом, способным к сложным движениям. Образование клюва и одновременно увеличение длины и подвижности шеи освободило передние конечности от многих добавочных функций (захват и удерживание добычи, чистка покровов и т. п.) и тем самым способствовало их превращению только в орган полета — крылья. Удлинение шеи и возрастание ее подвижности компенсировало неподвижность туловища и дало возможность, изменяя положение головы, в каких-то пределах менять положение центра тяжести тела, что важно при изменении режима полета. Преобразование передних конечностей в орган полета — крылья сопровождалось укреплением тазового пояса и скелета задних конечностей, а также возрастанием массы их мышц, обеспечивающих возможность «двуногого» движения на земле, по ветвям деревьев и т. п. Полет требует затраты большого количества энергии. Например у голубя, летящего со скоростью 70 км/ч, расход энергии в 27 раз больше, чем в покое. Резкое возрастание энергетических возможностей обеспечивается изменениями многих систем органов. Легкие птиц относительно малы по объему, но велика их губчатость (и тем самым большая поверхность окисления). Сложная система воздушных мешков, связанная с легкими, занимает до 20% объема тела — она резко увеличивает объем вдыхаемого воздуха и дает возможность насыщать кровь кислородом и во время вдоха, и во время выдоха. Чем интенсивнее полет, тем чаще дыхание1, что обеспечивает более интенсивное насыщение крови кислородом и удаление из нее углекислоты при ускорении движения. Одновременно воздушные мешки осуществляют интенсивную теплоотдачу во время полета, предохраняя организм летящей птицы от перегрева. Полное разделение большого и малого кругов кровообращения резко увеличивает возможность насыщения тканей кислородом. Сердце у птиц относительно очень велико — составляет 0,8—2,5% от веса тела (у рептилий 0,2—0,3%; у млекопитающих 0,2—1,5%). Относительно очень велика у птиц и частота сердцебиений: сердце крупных птиц сокращается 140—200 раз в 1 мин, мелких — до 500—600. В полете частота сердцебиений резко возрастает: так, у голубя в покое около 170 сердцебиений в 1 мин, а в полете 350—600; у мелких птиц частота пульса в полете до 1000 и более. Большой объем сердца и высокая частота сердцебиений создают быструю циркуляцию крови по организму, обеспечивая интенсивное насыщение тканей и органов питательными веществами и кислородом и освобождение их от углекислоты и продуктов распада. Дифференцировка желудка на два отдела — железистый и мускульный — обеспечивает механическую переработку пищи (в мускульном желудке может развиваться давление до 20—30 кг на 1 см2), что значительно увеличивает интенсивность ее переваривания. Этому же помогает свойственная ряду видов птиц способность отбрасывания погадок, т. е. удаления непереваренных остатков пищи (хитин, шерсть, кости и т. п.) из желудка через пищевод. Обеспечивает интенсивное пищеварение и высокая активность пищеварительных ферментов. Поэтому скорость переваривания пищи у птиц заметно выше, чем у пресмыкающихся и млекопитающих. Так, воробей переваривает гусеницу за 15 мин, жука — за 1 ч, зерно — за 3—4 ч (считая от времени принятия пищи до вывода ее непереваренных остатков). Интенсивное пищеварение позволяет перерабатывать большое количество пищи, обеспечивая высокие энергетические потребности организма. Особенности пищеварительной системы и органов захвата пищи наряду с большой подвижностью, высоким уровнем высшей нервной деятельности и совершенством органов чувств способствовали образованию широких пищевых связей и создали возможность использования очень большого числа кормовых объектов. Интенсивность пищеварения, кровообращения и дыхания обеспечивает высокий уровень обмена веществ, что наряду с образованием высоко эффективного теплоизолирующего покрова из пуха и перьев приводит к теплокровности птиц (гомойотермия), т. е. к их способности поддерживать температуру тела на постоянном уровне независимо от изменения температуры окружающей среды. Косвенным показателем высокого уровня обмена веществ птиц может служить температура их тела, которая у крупных видов равна 38—40°С, а у мелких видов достигает 41—43°С. Значительное увеличение объема головного мозга и возрастание числа нервных клеток в нем, по сравнению с рептилиями, определяют - более высокий уровень высшей нервной деятельности птиц. Это проявляется и в очень сложных системах врожденных рефлексов, определяющих поведение каждой особи, и в значительном возрастании роли индивидуального опыта, т. е. в выработке условных рефлексов. Плодовитость птиц меньше, чем у пресмыкающихся. Увеличение размеров яиц (откладка которых обеспечивается открытым тазом) увеличивает размеры вылупившегося детеныша. Сопутствующие размножению сложные биологические явления (постройка гнезда, насиживание, выкармливание, обогрев и охрана птенцов и т. п.) обеспечивают ускорение эмбрионального и постэмбрионального развития и повышенное выживание потомства. Совершенство органов движения (большая подвижность), гомойотермия, сложность высшей нервной деятельности и совершенство органов чувств (особенно зрения и слуха) позволили птицам широко расселиться по всему земному шару. Практически нет на земле ни одного района, включая высокие широты Арктики и Антарктики, где не встречались бы те или иные виды птиц. Более высокий уровень организации птиц, по сравнению с пресмыкающимися, находит свое отчетливое выражение не только в значительно более широком их распространении по всему земному шару, но и в более сложных и разнообразных взаимоотношениях со средой, с условиями жизни. Так, при сезонных неблагоприятных изменениях условий жизни (наступление зимы) пресмыкающиеся и земноводные прекращают свою активность, впадают в состояние оцепенения. У птиц в целом реакции при наступлении неблагоприятных сезонов года многообразнее и шире. Часть птиц совершает миграции (протяженностью до 5—10 тыс. км и более), улетая на зимовки в благоприятные по погодным и кормовым условиям районы. Однако значительное число видов остается в умеренных и северных широтах в течение всего года. Переживать суровые условия зимы им помогают довольно разнообразные приспособления. Для многих видов свойственна резко выраженная сезонная смена кормов: переход зимой на питание менее калорийными, но массовыми кормами (например, тетерев летом питается насекомыми, ягодами, зелеными частями растений, зимой — преимущественно почками и сережками некоторых пород деревьев). Неперелетные (оседлые) птицы совершают небольшие перемещения, концентрируясь в участках с большим количеством пищи (глухарь в сосновых лесах, где питается хвоей сосны; синицы, вороны — в населенных пунктах и т. п.). Некоторые виды осенью запасают пищу, пряча ее в укромных местах (кедровки, сойки, поползни — орехи лещины, «орешки» кедра, желуди; синицы — семена деревьев и кустарников). Оседлые птицы во время осенней линьки получают перьевой покров с лучшими теплоизолирующими свойствами, чем летний (большее развитие пуховой части пера, большее число перьев и пуха на единицу поверхности тела и т. п.); у некоторых видов зимний наряд резко отличается по окраске, помогая маскировке (белая куропатка). У части видов отчетливо выражены и приспособительные особенности поведения: ночевки в укрытиях (тетеревинные и некоторые другие птицы — в снегу; мелкие воробьиные — в дуплах, щелях за корой и т. п.). При этом птицы некоторых видов в сильные морозы ночуют группами, взаимно обогревая в убежище друг друга (синицы, воробьи и др.). Несмотря на резкое повышение уровня жизнедеятельности и широкие экологические возможности, открываемые гомойотермией, отчетливая специализация птиц к полету в известной мере ограничивает круг экологических приспособлений этих животных. Отсутствие столь узкой специализации у млекопитающих создало предпосылки к возникновению в процессе эволюции этого класса значительно большего разнообразия жизненных форм и более многообразных взаимоотношений со средой. Дополнительная литература Гладков Н. А., Дементьев Г. П., Птушенко Е. С., Судиловская А. М. Определитель птиц СССР. М., 1964. Гуртовой Н. Н., Матвеев Б. С, Дзержинский Ф. Я. Практическая зоотомия позвоночных. Птицы, млекопитающие. М., 1980. Дементьев Г. П. Птицы. Руководство по зоологии, т. 6. М., 1940. Дементьев Г. П. Птицы нашей страны. М., 1962. Иванов А. И., Штегман Б. К. Краткий определитель птиц СССР. Л., 1964. Карташев Н. Н. Систематика птиц. М., 1974. Михеев А. В. Биология птиц. М., 1960. Шмальгаузен И. И. Основы сравнительной анатомии. М., 1947. Шульпин Л. М. Орнитология. Л., 1940. ТЕМА 18. ОПРЕДЕЛЕНИЕ ПТИЦ МАТЕРИАЛ И ОБОРУДОВАНИЕ Комплект коллекционного материала (тушки 40—50 видов птиц). На двух — четырех студентов необходимы: 1. Линейка длиной 50 см или обычная сантиметровая лента — 1. 2. Штангенциркуль или циркуль — 1. ЗАДАНИЕ 1. Определить по одному представителю всех имеющихся в наборе отрядов птиц; кратко записать основные, «ключевые» признаки каждого отряда. 2. По указанию преподавателя, часть птиц определить до вида (последовательно определяя отряд, семейство, род и вид). 3. Обратить внимание на связь особенностей внешнего облика с образом жизни (приспособления к водному образу жизни, ловле мелкой добычи в воздухе и т. д. и т. п.). Распределить определенные виды по их образу жизни (водные птицы, околоводные, ловящие добычу в воздухе, собирающие ее на стволах деревьев и т. п.). 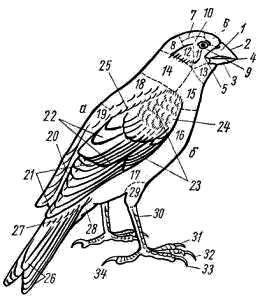 Рис. 113. Названия участков оперения и частей тела птицы. а — спинная, или верхняя, сторона тела; б — брюшная, или нижняя, сторона тела: 1 — надклювье, 2 — гребень, или конек, надклювья, 3 — подклювье, 4 — вершина клюва, 5 — угол рта, 6 — лоб, 7 — темя, 8 — затылок, 9 — уздечка, 10 — бровь, 11 — щека, 12 — кроющие перья уха, 13 — горло, 14 — шея, 15 — зоб, 16 — грудь, 17 — брюхо, 18 — передняя часть спины, 19 — задняя часть спины, 20 — поясница, 21 — первостепенные маховые, 22 — второстепенные маховые, 23 — верхние кроющие крыла, 24 — кроющие кисти, 25 — плечевые перья, 26 — рулевые перья, 27 — верхние кроющие хвоста, 28 — нижние кроющие хвоста, 29 — голень, 30 — цевка, 31 — внутренний палец, 32 — средний палец, 33 — наружный палец, 34 — задний палец Перед определением по рисункам (рис. 113, 114) ознакомиться с обозначениями отдельных участков оперения и принятыми в систематике измерениями частей тела птиц. 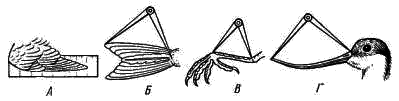 Рис. 114. Измерение длины частей теля птицы. А — длина крыла; Б —длина хвоста; В — длина цевки; Г — длина клюва ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ И ПОДОТРЯДОВ ПТИЦ . 1(4). Края надклювья и подклювья усажены поперечными пластинками или хорошо развитыми роговыми зубчиками (рис.115) См. 2 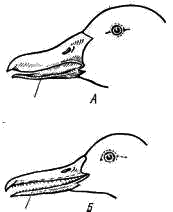 Рис. 115. Клюв. А — кряквы (края надклювья и подклювья несут поперечные роговые пластинки); Б — длинноклювого крохаля (края надклювья и подклювья несут роговые зубчики) 2(3). Цевка длиннее 200 мм Отряд Аистообразные, или Голенастые, Ciconiiformes . 3(2). Цевка короче 200 мм Отряд Гусеобразные, Anseriformes . 4(1). Края надклювья и подклювья без пластинок и зубцов (хотя могут быть слегка зазубренными) См. 5 . 5(14). Пальцы соединены плавательной перепонкой (рис. 116, 118). Передний ее край может быть более или менее глубоко вырезан, но в этом случае перепонка между средним и внутренним пальцами не короче половины длины внутреннего пальца См. 6 . 6(7). Все 4 пальца соединены плавательной перепонкой (рис. 116) Отряд Пеликанообразные, или Веслоногие, Pelecaniformes |