Происхождение и филогения простейших.. Зоология.. Происхождение и филогения простейших
 Скачать 1.17 Mb. Скачать 1.17 Mb.
|

|
Происхождение и филогения простейших. Простейшие (Protozoa) относятся к примитивным одноклеточным эукариотам (надцарство Eucaryota). В настоящее время общепризнано, что эукариоты произошли от прокариот. Об их единстве свидетельствует сходство процессов синтеза белка в клетке. Прокариоты развились на планете раньше эукариот, это подтверждают ископаемые остатки их жизнедеятельности, а также способность ряда прокариот существовать в бескислородной среде (как доказано, атмосфера Земли около 2 млрд. лет назад была восстановительной). Существуют две гипотезы происхождения эукариот от прокариот. Сукцессивная гипотеза утверждает, что мембранные органеллы клетки (ядро, митохондрии, пластиды, аппарат Гольджи) возникли постепенно (сукцессивно) из мембраны клетки прокариот. Эндосимбиотическая гипотеза предполагает, что в эволюции эукариот большую роль сыграл симбиоз различных прокариот. Допускается, что митохондрии и хлоропласты могли развиться из симбиотических бактерий, живших в клетке хозяина. Однако каждая из указанных гипотез имеет слабые стороны. Пока нет убедительных доказательств эволюции эукариот, что затрудняет выяснение эволюционных взаимоотношений между современными Protozoa. Из известных нам семи типов простейших четыре (Apicomplexa, Myxozoa, Microspora, Ascetospora) исключительно паразитические группы, возникшие явно поздно, после появления в процессе эволюции их хозяев — высших многоклеточных. Поэтому при выяснении наиболее примитивной группы среди простейших, которую можно было бы сопоставить с предковой, следует обратиться к свободноживущим формам, к которым относятся три типа. Из них инфузории относятся к самым высокоорганизованным простейшим, многие из которых обладают надклеточным уровнем организации и являются полиэнергидными. Лабиринтулы несут черты многоклеточности и чрезвычайно специализированы. Только саркомастигофоры обладают многими первичными признаками (плезиоморфными), общими с предками всех простейших. Однако саркомастигофоры неоднородны, и особенно резко различаются среди них подтипы Sarcodina и Mastigophora. Мысль о том, какая из этих групп простейших ближе к предкам, занимала и занимает умы многих современных ученых. Так, Пашер (1914) предположил, что наиболее плезиоморфной группой следует считать жгутиковых. У жгутиковых близкая связь с одноклеточными растениями, разнообразные типы питания, а органеллами движения являются жгутики, которые встречаются даже у прокариот. Кроме того, жгутики имеются у гамет Protozoa и Metazoa. Упрощенность организации саркодовых (отсутствие жгутиков, пелликулы) Пашер считал вторичным явлением в связи с переходом к активному фагоцитозу (анимальному способу питания), а наличие жгутиковых гамет у некоторых саркодовых рассматривал как свидетельство их происхождения от жгутиковых. Ряд других ученых, в том числе и отечественный ученый А. Н. Опарин (1924), придерживались другой позиции и рассматривали в качестве первичной группы саркодовых. По их мнению, отсутствие оболочки, непостоянство формы тела, наличие псевдоподий и гетеротрофное питание — примитивные признаки. Это соответствовало постулатам популярной в то время теории происхождения жизни на Земле А. Н. Опарина. Согласно этой теории, образование первых существ произошло от белковых комочков (коацерватов), похожих на амеб. Опарин признавал первичность гетеротрофов, питавшихся готовыми органическими веществами в первичном «бульоне» океанов. В дальнейшем выяснились новые факты, проливающие свет на происхождение Protozoa. Во-первых, среди Procaryota обнаружено восемь типов метаболизма, поэтому вопрос о том, какой способ питания был у первичных одноклеточных (Eucaryota), усложнился. А изучение трофики Mastigophora показало, что во многих группах зеленых жгутиконосцев (Phytomastigophorea) наблюдается утрата автотрофности и переход к гетеротрофности. По-видимому, разнообразие способов питания у жгутиковых явилось основой для дальнейшей их дивергенции на автотрофов и гетеротрофов. Появились и новые доказательства вторичной упрощенности саркодовых. Так, у некоторых амеб обнаружены отстатки кинетосомы в цитоплазме, что свидетельствует о редукции жгутиков у взрослых форм. А с другой стороны, описано немало случаев модификации клеток со жгутиками в амебоидные (у колониальных форм и многоклеточных). Кроме того, имеются формы, обладающие псевдоподиями и жгутиками одновременно. На основании этих данных подавляющее большинство ученых придерживаются точки зрения Пашера и предполагают, что предками современных Protozoa были древние Sarcomastigophora с разнообразными способами питания и со жгутиками примитивного строения. От этих предковых форм развились три ветви: саркодовые и эволюционно продвинутые жгутиконосцы, опалиновые. В настоящее время не вызывает сомнений родство споровиков (Apicomplexa) и инфузорий со жгутиковыми. Первые упростились в строении, по-видимому, в связи с паразитизмом, но при этом усложнили свой жизненный цикл. Однако у них сохранились жгутиковые гаметы и зиготическая редукция хромосом, что подтверждает их родство со жгутиковыми. У инфузорий произошла полимеризация многих органелл и возникла особая форма полового процесса — конъюгация. Такие типы, как Myxozoa и Microspora, возможно, произошли от каких-то древних саркодовых, так как у них развитие начинается с амебоидного зародыша и жгутиковых гамет нет. Однако жизненный цикл и образование спор у этих типов не находят аналогов среди Protozoa, и поэтому некоторые авторы признают автономность их эволюционного развития. Своеобразны и такие типы, как лабиринтулы и асцетоспоридии, у которых не прослеживаются черты сходства с другими группами. Филогения 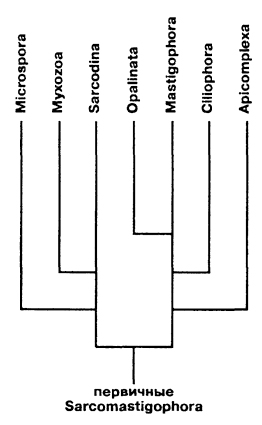 Теории происхождения многоклеточных животных. Теории Мечникова и Геккеля. Теории происхождения многоклеточных животных. Теории Бючли и Иванова. Русский биолог И.И. Мечников в 1882 г. опубликовал другую колониальную гипотезу — «фагоцителлы», раскрывающую сущность происхождения многоклеточных. Гипотеза базировалась на обширных исследованиях автора. Мечников открыл явление фагоцитоза — внутриклеточного пищеварения у многоклеточных и считал этот способ переваривания пищи более примитивным, чем полостное пищеварение. По его мнению, первые многоклеточные были примитивнее «гастреи» по организации и не имели еще пищеварительной полости и полостного пищеварения. Для выяснения вопроса о гипотетическом предке Metazoa И. И. Мечников пристально изучал онтогенез примитивных многолеточных — губок. Им было обнаружено, что образование двуслойной фазы развития у губок происходит не путем инвагинации бластулы, а путем иммиграции отдельных клеток наружного слоя в полость зародыша (бластоцель). Личинка губок с паренхимными клетками внутри была названа паренхимулой. И. И. Мечников рассматривал паренхимулу как прообраз или живую модель гипотетического предка многоклеточных — фагоцителлы 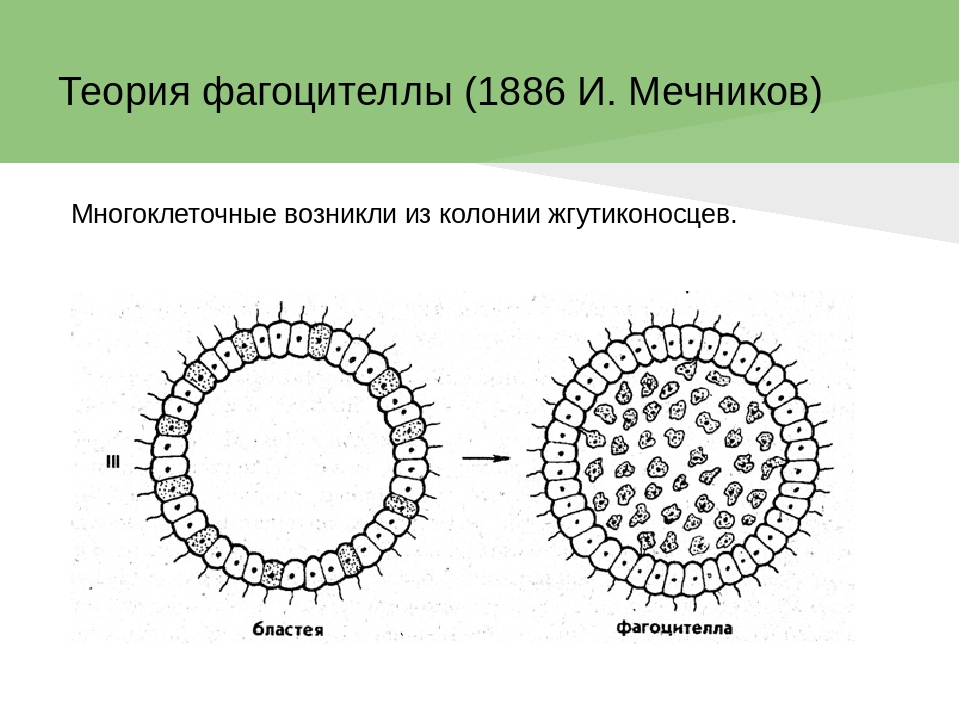 Это название предка связано со способом питания — фагоцитозом, который осуществлялся в паренхиматозных клетках. По его мнению, фагоцителла могла возникнуть из шаровидных колоний жгутиконосцев путем иммиграции части клеток внутрь колонии. При этом наружные клетки со жгутиками выполняли функцию движения (кинобласт), а внутренние — утрачивали жгутики, становились амебоидными и выполняли функцию фагоцитоза (фагоцитобласт). Гипотеза фагоцителлы И. И. Мечникова завоевала широкое признание и нашла дальнейшее развитие в трудах многих ученых. Дополнения и существенные поправки к теории фагоцителлы внесли советские ученые А. А. Захваткин и А. В. Иванов. Так, А. А. Захваткин (1949) создал гипотезу синзооспоры. По его мнению, нельзя было принимать в каче- :тве предка многоклеточных колонии зеленых жгутиконосцев (типа Volvох), как это допускали предшественники, так как у вольвоксовых голофитный способ питания и зиготическая редукция хромосом, как у растений. Поэтому А. А. Захваткин предположил, что колониальные простейшие, давшие начало Metazoa, обладали не голофитным, а голозойным типом питания и имели гаметическую редукцию хромосом в онтогенезе. 3 связи с тем, что у всех Metazoa эмбриогенез протекает в пределах яйцевой оболочки и дробление зародыша вначале палинтомическое и только после выхода зародыша из яйца дробление становится монотомическим, Захваткин предположил, что колониальный предок в своем индивидуальном развитии также проходил подобные фазы дробления. Другая важная поправка касалась облика первого многоклеточного животного. Захваткин считал, что фагоцителла И. И. Мечникова отражает облик не взрослого предка многоклеточных, а лишь его личинки — синзооспоры. А взрослая фаза предка многоклеточных, по мнению Захваткина, представляла сидячую форму колониального типа, похожую на губок. Но гипотеза синзооспоры не получила широкого распространения, так как трудно было допустить, чтобы сидячие колониальные формы могли дать дальнейшую эволюцию всех Metazoa. Первую колониальную гипотезу происхождения Metazoa разработал зоолог-эволюционист Э. Геккель (1874), гипотеза получила название «гастреи» (рис. 67, I). Он считал, что протозойным предком Metazoa была «бластея» — шаровидная колония жгутиковых, похожая на стадию бластулы в развитии многих многоклеточных. В процессе эволюции от бластеи путем инвагинации (впячивания) могли возникнуть первые двуслойные многоклеточные с кишечной полостью, выстланной энтодермой. Этот гипотетический предок Metazoa был назван Э. Геккелем «гастреей» в связи со сходством со стадией гаструлы в развитии многоклеточных. Гастрея, по Геккелю, представляла плавающее двуслойное животное со ртом. Наружный слой жгутиковых клеток гастреи представлял эктодерму и выполнял двигательную функцию, а внутренний слой клеток (энтодерма) — пищеварительную. От гастреи, по его мнению, произошли прежде всего двуслойные животные — кишечнополостные. 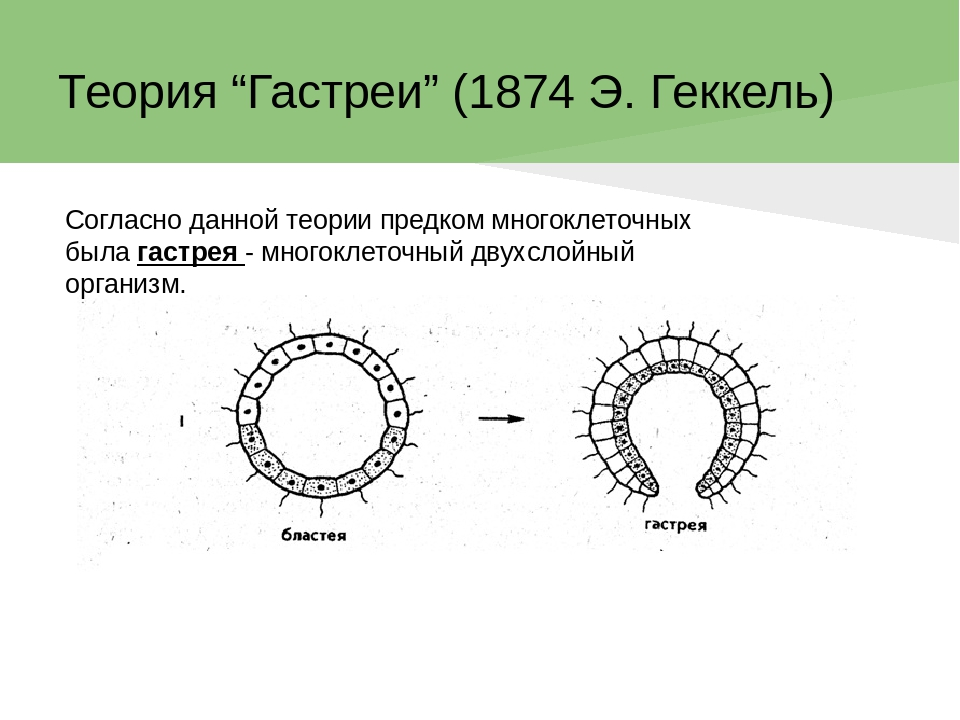 Дальнейшее развитие теории «гастреи» продолжил О. Бючли (1884), предложивший ее новый вариант — гипотезу «плакулы» (рис. 67, II). По правка Бючли к теоретическим рассуждениям Геккеля состояла в том, что он считал колониальных простейших типа «бластеи» эволюционно продвинутыми и предложил в качестве гипотетического колониального предка более простую пластинчатую колонию одноклеточных типа современных Gonium Путем расщепления такой пластинки на два слоя возник, по Бючли, гипотетический предок многоклеточных — «плакула». В дальнейшем из плакулы могла образоваться, по Бючли, гастрея путем чашевидного прогибания двуслойной пластинки. Натолкнуло Бючли на создание нового варианта гипотезы описание примитивного двуслойного многоклеточного животного — трихоплакса (Trichoplax), строение которого приближалось к плоским колониям жгутиковых. Автор предполагал, что подобные трихоплаксу животные могли быть промежуточными между «плакулой» и «гастреей». Тем более было обнаружено, что у Trichoplax нижний слой клеток способен к наружному пищеварению. Ползая, эти животные выделяют пищеварительные соки (ферменты), переваривающие бактериальную пленку. В дальнейшей эволюции, по его мнению, из этого нижнего слоя клеток примитивных плакулоподобных организмов возникла энтодерма гастреи, а из верхнего — эктодерма. 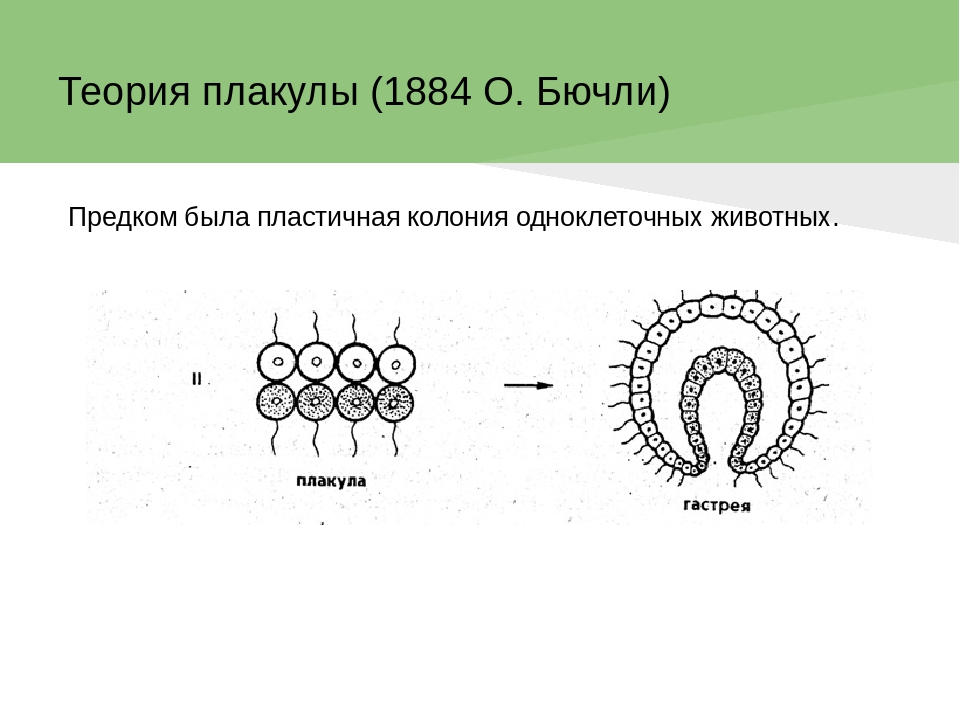 Крупный современный зоолог А. В. Иванов (1967) синтезировал современные идеи по проблеме происхождения многоклеточных. За основу он принял гипотезу фагоцителлы Мечникова. Однако он предложил считать в качестве колониального предка Metazoa колонию типа воротничковых жгутиковых, имеющих голозойный способ питания, что соответствовало взглядам А. А. Захваткина. Живой моделью фагоцителлы А. В. Иванов считает не столько личинку губок — паренхимулу, сколько трихоплакса, близкого по организации к фагоцителле. Фагоцителла, по А.В.Иванову, в процессе эволюции дала начало таким типам, как Губки (Spongia) и Пластинчатые (Placozoa), обладающим примитивным внутриклеточным пищеварением — фагоцитозом. Согласно взглядам А. В. Иванова, появление двуслойных животных со ртом, кишечной полостью и полостным пищеварением произошло значительно позднее, чем фагоцителлоподобных . 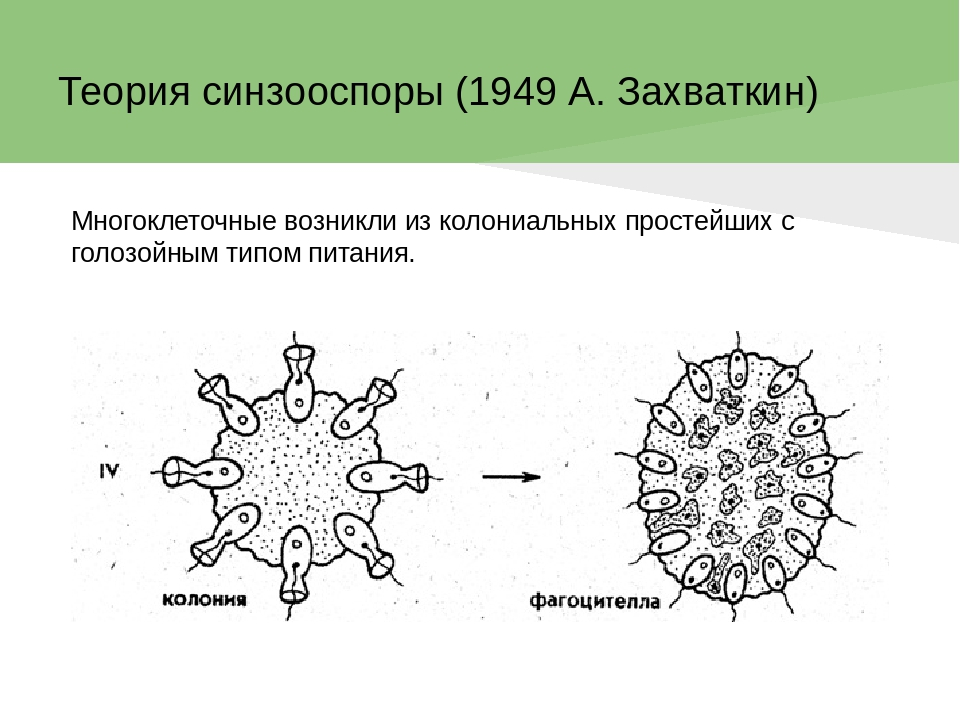 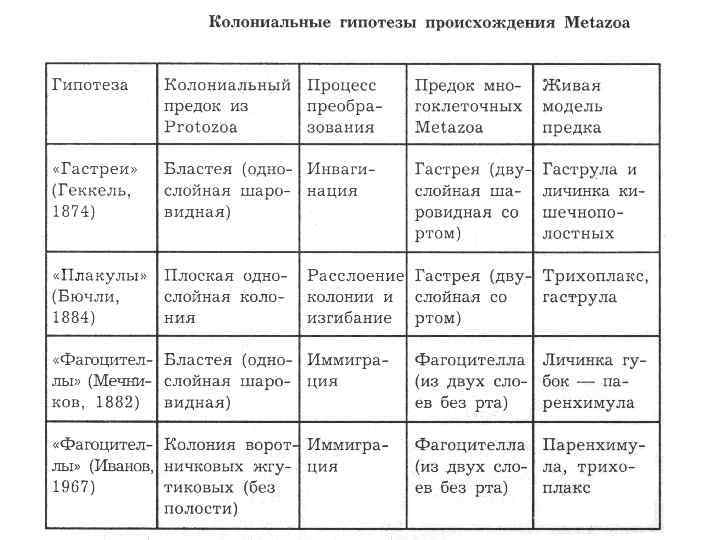 Трихоплакс и его место в животном мире. К типу пластинчатых относятся всего лишь два вида одного рода морских животных — Trichoplax. Это мелкие плоские животные, движущиеся по водорослям при помощи покровных жгутиковых клеток. Внутри тела трихоплакса имеются фагоцитарные клетки с пищеварительными вакуолями .История открытия типа необычна. Немецкий зоолог Ф. Шульце в 1883 г. описал новый вид примитивного многоклеточного животного, обнаруженного в морском аквариуме, и назвал его Trichoplax adhaerens. Hо сведения о его биологии и развитии были отрывочны и недостаточны. В начале XX в. Т. Крумбах (1907) предположил, что Trichoplax — личинка медузы, и потому интерес к этому необычному объекту угас. Только в 1971 г. паразитолог К. Грелль обнаружил, что Trichoplax размножается половым путем и, следовательно, представляет собой взрослый организм. Ему удалось наблюдать образование яйцеклеток и их дробление. Так был открыт новый тип самых примитивных реликтовых многоклеточных — Placozoa. Советский зоолог А. В. Иванов (1973) рассматривает трихоплакса как живую модель для современной теории происхождения многоклеточных, а тип пластинчатых им выделен в особый надраздел — Phagocytellozoa. Трихоплакс — пластинкообразное морское животное, размером не более 4 мм. Обитает на водорослях. Внешне напоминает амебу, так как не имеет постоянной формы тела и при движении его контуры меняются (рис. 69, А). Однако движется он при помощи жгутиковых клеток, покрывающих тело. Жгутиковые клетки «брюшной» стороны узкие и высокие, а на «спинной» поверхности — уплощенные. Внутри тела рыхло расположены веретеновидные и амебоидные клетки . Питание трихоплакса происходит двумя способами. К. Грелль описал способ внешнего пищеварения у трихоплакса. Он обнаружил, что клетки "брюшной» стороны способны выделять ферменты и переваривать пленку бактерий, покрывающую субстрат. К. Грелль считал, что это единственный способ питания у трихоплакса, и потому придерживался гипотезы происхождения пластинчатых от «плакулы» (по О. Бючли), согласно которой предок многоклеточных был двуслойной колонией жгутиконосцев. Второй способ питания трихоплакса — фагоцитоз. Пищеварительные вакуоли в клетках трихоплакса были обнаружены давно, но их природа не была доказана. Только в 1986 г. немецкий ученый Вендерот экспериментально доказал фагоцитоз у трихоплакса. Он кормил трихоплаксов убитыми дрожжевыми клетками и выяснил, что фагоцитоз для них основной способ питания. Трихоплаксы загоняли пищевые частицы биением жгутиков покровных клеток на спину, где пища поглощалась веретеновидными клетками, которые способны выступать наружу через поры между жгутиковыми клетками. Перегруженные пищеварительными вакуолями клетки становились короткими, амебоидными и занимали свое место в толще паренхимы. После переваривания пищи они снова вытягивались и становились веретеновидными. Так подтвердилась идея И. И. Мечникова о фагоцитозе как наиболее примитивном способе питания у многоклеточных. Размножение трихоплакса, как показал К. Грелль, происходит бесполым и половым путем. Бесполое размножение осуществляется делением тела надвое или путем отшнуровывания «бродяжек» со спинной стороны. Известен и половой процесс у трихоплакса. Если яйцеклетки были обнаружены Греллем в 1971 г., то спермин в форме S-клетки им были выявлены только в 1981 г. Выяснено, что сперматозоиды у трихоплаксов без жгутиков. Оплодотворенные яйцеклетки претерпевают равномерное дробление, которое прослежено до 32 бластомеров. Личинка трихоплакса пока неизвестна. Клетки трихоплакса не полностью утратили самостоятельность. При извлечении из тела они сползаются в агрегаты и могут постепенно развиваться в целый организм. Клетки трихоплакса обладают высокими регенерационными возможностями. Всего описано два вида трихоплаксов из Атлантического и Тихого океанов. Они хорошо содержатся в морских аквариумах и являются прекрасными лабораторными объектами. Анализ морфологии трихоплакса дает основание утверждать, что это действительно фагоцителлообразный организм, по-видимому, потомок реликтовой фауны первых многоклеточных животных на Земле. У трихоплаксов нет еще зародышевых листков, кишечника, рта, нервной системы и мышечных клеток. Несомненно, что это самые примитивные современные многоклеточные. Губки Губки — неподвижные прикрепленные животные, обитающие преимущественно в морях, реже в пресных водах. Они имеют форму наростов, ковриг, бокалов или напоминают ветвящиеся стебли . Губки могут быть одиночными животными, но значительно чаще образуют колонии. Долгое время губки относили к зоофитам — промежуточным формам между растениями и животными. Принадлежность губок к животным впервые была доказана Р. Эллисом в 1765 г., который обнаружил явление фильтрации воды через тело губок и голозойный тип питания. Р. Грант (1836) впервые выделил губок в самостоятельный тип Губки (Porifera). Всего известно 5000 видов губок. Это древняя группа животных, известная с докембрия. Общая характеристика типа губок. Губки сочетают в себе признаки примитивных многоклеточных животных со специализацией к неподвижному образу жизни. О примитивности организации губок свидетельствуют такие признаки, как отсутствие тканей, органов, высокая регенерационная способность и взаимопревращаемость многих клеток, отсутствие нервных и мышечных клеток. Им свойственно только внутриклеточное пищеварение. С другой стороны, губки несут черты специализации к неподвижному образу жизни. У них имеется скелет, защищающий тело от механических повреждений и хищников. Скелет может быть минеральный, роговой или смешанной природы. Обязательным компонентом скелета является роговое вещество — спонгин (отсюда одно из названий типа — Spongia). Тело пронизано порами. Это отражено в синониме названия типа — Porifera (pori — поры, fera — несущие). Через поры вода поступает внутрь тела со взвешенными пищевыми частицами. С током воды через тело губок пассивно осуществляются все функции: питание, дыхание, выделение, размножение. В процессе онтогенеза происходит извращение (инверсия) зародышевых пластов, т. е. первичный наружный слой клеток занимает положение внутреннего слоя, и наоборот. Выделяют три класса губок: класс Известковые губки (Calcispongiae), класс Стеклянные губки (Hyalospongiae), класс Обыкновенные губки (Demospongiae). У губок различают следующие основные типы клеток: Пинакоциты — плоские многоугольные, способные сокращаться клетки, которые выстилают наружную поверхность и поровые каналы Г.. Воротничкоые клетки, или хоаноциты, цилиндрической или шаровидной формы. Они образуют жгутиковые камеры, а у наиболее примитивных одиночных известковых губок выстилают внутреннюю (гастральную) полость. На свободном конце клетки, обращенном внутрь жгутиковой камеры, расположен жгутик, окруженный цитоплазматическим воротничком (рис. 2). С помощью электронного микроскопа выяснено, что воротничок хоаноцита образован венчиком из 30—40 тесно прилежащих друг к другу тончайших плазматических выростов — щупалец. Жгутик хоаноцита отходит от базального тельца, к которому присоединено чешуевидное парабазальное тельце (рис. 2, В). Такое же ультратонкое строение имеют и воротничковые жгутиковые (Craspedomonadina), что указывает на их филогенетическую близость. Полагают, что Г. и воротничковые жгутиковые имели общего предка. Хоаноциты выполняют вододвигательную и питательную функцию. Биение их жгутиков создает постоянный направленный ток воды в теле Г., а пищевые частицы, приносимые водой, захватываются и перевариваются хоаноцитами. Весь процесс пищеварения у губок проходит внутриклеточно. Непереваренные остатки пищи из пищеварительных вакуолей выбрасываются с током воды через оскулум. Амебоциты — крупные блуждающие клетки, они осуществляют фагоцитоз и участвуют в переваривании пищи, получая излишек пищевых частиц от хоапоцитов. Амебоциты постоянно передвигаются в студенистой массе основного вещества Г. — мезоглее, которая заполняет все пространство между поверхностью Г. и системой водоносных каналов. Мезоглея связывает все типы клеток механически и химически, или гуморальн Колленциты — звездчатые клетки, соприкасающиеся своими отростками с соседними клетками, они выполняют опорную функцию. Склеробласты и сионгиобласты — крупные клетки, формирующие элементы скелета Г. — спикулы и спонгинотше волокна. Археоциты — не дифференцированные клетки, способные легко превращаться в клетки любого типа, служат клеточным резервом. Половые клетки, образующиеся из археоцитов, обеспечивают половое размножение. Считается, что губки возникли в докембрии около 1 миллиарда лет назад. Расцвет данной группы живых организмов приходится на мезозой. Губки представлены в основном морскими организмами, лишь их небольшая часть относится к пресноводным. Они ведут неподвижный образ жизни, прикрепляясь к субстрату, практически не реагируя на раздражение. Чаще всего губки ведут колониальный образ жизни, иногда встречаются единичные особи. Губки имеют твердое и жесткое тело, часто обладают яркой окраской (за исключением серых и зеленоватых бадяг). Окраска их варьирует в зависимости от количества пигмента. Большое количество губок имеют неприятный запах и вкус. При всем примитивизме губок, их видовое разнообразие достаточно широкое. Тип включает в себя более 8000 видов. Если говорить о месте обитания губок, то в Черное море насчитывается их 300 видов, а Каспийском море - 1 вид. . Одиночные губки в про - стейшем случае имеют форму бо - кала, например Sycon (рис. 70, 2). Такая форма обладает гетеропо - лярной осевой симметрией. У бока - ловидной губки различают подош - ву, которой она прикрепляется к субстрату, а на верхнем полюсе — устье — оскулюм . Через тело губок постоянно осу - ществляется ток воды: через поры вода поступает в губку, а из устья выходит. Направление тока воды в губке определяется движением жгутиков особых воротничковых клеток. У колониальных губок име - ется множество устьев (оскулюмов) и осевая симметрия нарушается. Стенка тела губок состоит из двух слоев клеток: покров - ных клеток (пинакоцитов) и внут - реннего слоя жгутиковых ворот - ничковых клеток (хоаноцитов), ко - торые выполняют функцию филь - трации воды и фагоцитоза. Хоано - циты имеют вокруг жгутика ворот - ничок в форме воронки. Воротничок образован из сцепленных микро - ворсинок. Между слоями клеток имеется студенистое вещество — мезоглея, в которой расположены отдельные клеточные элементы. К ним относятся: звездчатые опорные клетки (колленциты), скелетные клетки (склероциты), подвижные амебоидные клетки (амебоциты) и недифференцированные клетки — археоциты, которые могут давать начало любым другим клеткам, в том числе и половым. Иногда присутствуют слабосокращающиеся клетки — миоциты. Среди пинакоцитов различают особые клетки — пороциты со сквозной порой. Пороцит способен к сокращению и может открывать и закрывать пору. Поры рассеяны по всему телу губки или образуют скопления. Различают три типа морфологического строения губок: аскон, сикон, лейкон (рис. 72). Наиболее простой из них аскон. Асконоидные губки — мелкие одиночные, у которых вода поступает через поры и поровые каналы, пронизывающие стенку тела, в атриальную полость, выстланную хоаноцитами, а затем через оскулюм выходит наружу. Губки типа сикон крупнее, с более толстыми стенками, в которых имеются жгутиковые камеры. Ток воды в губках сиконоидного типа происходит по следующему пути: поры, поровые каналы, жгутиковые камеры, атриальная полость, оскулюм. В отличие от асконоидных губок у сиконоидных хоаноциты выстилают не атриальную полость, а многочисленные жгутиковые карманы в толще стенки тела. Это увеличивает пищеварительную поверхность губок и повышает эффективность фагоцитоза. Атриальная полость у сиконоидных выстлана пинакоцитами. Наиболее сложный тип строения — лейкон. Это колониальные губки с многочисленными оскулюмами. В мощном слое мезоглеи много скелетных элементов. Стенка тела пронизана сетью каналов, связывающих многочисленные жгутиковые камеры. Ток воды в лейконоидной губке осуществляется по пути: поры — поровые каналы — жгутиковые камеры — выносящие каналы — атриальная полость — оскулюм. У лейконоидных губок наибольшая пищеварительная поверхность. Тип строения губок не отражает их систематического родства. В разных классах губок имеются представители с разным морфологическим строением. Это свидетельствует о параллельных путях эволюции в разных классах губок. Преимущество усложнения строения губок оказалось в том, что с увеличением размеров тела губок увеличивалась пищеварительная поверхность слоя хоаноцитов и усиливалась интенсивность фильтрации. Например, губка Leuconia (лейкон) размером в 7 см профильтровывает за сутки 22 л воды |