Коллоквиум по зоологии беспозвоночных. Губки. Коллоквиум. Губки. Особенности организации губок как низших многоклеточных существ
 Скачать 212.5 Kb. Скачать 212.5 Kb.
|

|
Особенности организации губок как низших многоклеточных существ Губки - неподвижные прикрепленные животные, обитающие преимущественно в морях реже пресных водах. Губки могут быть одиночными животными, но значительно чаще образует колонии. Теперь о самых общих признаках губок: Губки – это неподвижные животные. Губки являются примитивными многоклеточными животными, которые не имеют пищеварительной, кровеносной и нервной систем. Также у них нет органов, тканей, и клетки не организовывают четко определенную структуру. Вместо этого различные функции выполняют разнообразные отдельные клетки и клеточные пласты. (Например, амебоциты - подвижные клетки, которые осуществляют переваривания пищи и разнесения питательных веществ по телу губки). Также у губок отсутствуют нервные и мышечные клетки. Скелет губок — продукция клеток-склеробластов (клетки, внутри которых закладываются и развиваются отдельные скелетные элементы губок). Скелет состоит из отдельных мелких элементов — иголок (спикул (элементы скелета, выполняющие опорную функцию и поддерживающие форму тела губок)). Эти иголочки имеют самую различную форму: звездчатые, в форме снежинок и кристалликов. Состоят спикулы из кремнезема, извести либо из органического вещества спонгина, напоминающего роговое вещество. Наличие пор в стенках тела губок. Через них в губку проникает вода с растворённым в ней кислородом – так происходит пассивное дыхание губок. У губок хорошо развита регенерация. Губки легко регенерируют поврежденные участки тела – рана зарубцовывается, затягивается защитной мембраной, под которой восстанавливается прежняя структура, и через некоторое время место повреждения становится незаметным. Для губок характерно внутриклеточное переваривание пищи. (Сквозь поры тела вместе с водой в губку попадают плавающие мелкие организмы, которые губка использует в пищу. Жгутиковые клетки, из которых построена энтодерма, захватывают их путем фагоцитоза и переваривают. Кроме того, специальные подвижные амебовидные клетки внутреннего слоя также захватывают пищу и переваривают ее) Характерны хоаноциты (обеспечивают циркуляцию воды и питание). Морфологические типы губок Наиболее просто организованные губки асконоидного типа имеют форму мешка, который основанием прикреплен к субстрату, а устьем (оскулумом) обращен кверху. Наружный слой стенки мешка образован покровными клетками (пинакоцитами), внутренний – воротничковыми жгутиковыми клетками (хоаноцитами). Хоаноциты выполняют функцию фильтрации воды и фагоцитоза. Между наружным и внутренним слоями располагается бесструктурная масса – мезоглея, в которой находятся многочисленные клетки, в том числе образующие спикулы (иглы внутреннего скелета). Все тело губки пронизано тонкими каналами, ведущими в центральную атриальную полость. Непрерывная работа жгутиков хоаноцитов создает ток воды: поры → поровые каналы → атриальная полость → оскулум. Питается губка теми пищевыми частицами, которые приносит вода. У губок сиконоидного типа происходит утолщение мезоглеи и образование внутренних впячиваний, имеющих вид карманов, выстланных жгутиковыми клетками. Ток воды в сиконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые карманы → атриальная полость → оскулум. Н 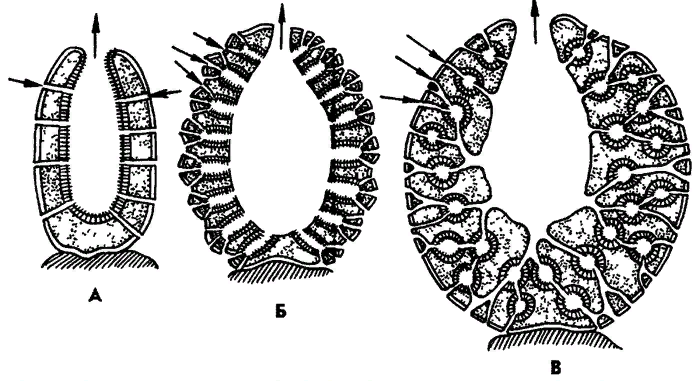 аиболее сложный тип губок – лейкон. Для губок этого типа характерен мощный слой мезоглеи с множеством скелетных элементов. Внутренние впячивания погружаются вглубь мезоглеи и приобретают вид жгутиковых камер, соединяющихся выносящими каналами сатриальной полостью. Атриальная полость у лейконоидных губок, так же как у сиконоидных выстлана пинакоцитами. Лейконоидные губки обычно образуют колонии с множеством устьев на поверхности: в виде корок, пластинок, комьев, кустов. Ток воды в лейконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые камеры → выносящие каналы → атриальная полость → оскулум. аиболее сложный тип губок – лейкон. Для губок этого типа характерен мощный слой мезоглеи с множеством скелетных элементов. Внутренние впячивания погружаются вглубь мезоглеи и приобретают вид жгутиковых камер, соединяющихся выносящими каналами сатриальной полостью. Атриальная полость у лейконоидных губок, так же как у сиконоидных выстлана пинакоцитами. Лейконоидные губки обычно образуют колонии с множеством устьев на поверхности: в виде корок, пластинок, комьев, кустов. Ток воды в лейконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые камеры → выносящие каналы → атриальная полость → оскулум.Особенности размножения и развития губок Р 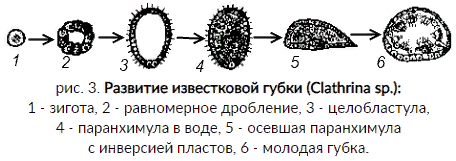 азмножаются бесполым и половым способами. Бесполое размножение осуществляется в форме наружного почкования, внутреннего почкования, фрагментации, образования геммул и пр. При половом размножении из оплодотворенной яйцеклетки развивается бластула, состоящая из одного слоя клеток, имеющих жгутики. Затем часть клеток мигрирует внутрь и превращается в амебоидные клетки. После того, как личинка оседает на дно, происходит перемещение жгутиковых клеток внутрь, они становятся хоаноцитами, а амебоидные клетки выходят на поверхность и превращаются в пинакоциты. азмножаются бесполым и половым способами. Бесполое размножение осуществляется в форме наружного почкования, внутреннего почкования, фрагментации, образования геммул и пр. При половом размножении из оплодотворенной яйцеклетки развивается бластула, состоящая из одного слоя клеток, имеющих жгутики. Затем часть клеток мигрирует внутрь и превращается в амебоидные клетки. После того, как личинка оседает на дно, происходит перемещение жгутиковых клеток внутрь, они становятся хоаноцитами, а амебоидные клетки выходят на поверхность и превращаются в пинакоциты.Далее личинка превращается в молодую губку. То есть первичная эктодерма (мелкие жгутиковые клетки) становится на место энтодермы, а энтодерма – на место эктодермы: зародышевые пласты меняются местами. На этом основании зоологи называют губок животными, вывернутыми наизнанку. Личинка большинства губок – паренхимула, по строению почти полностью соответствует гипотетической «фагоцителле» И.И. Мечникова. В связи с этим в настоящее время наиболее обоснованной считается гипотеза происхождения губок от фагоцителлообразного предка. Особенности организации и биологии трихопласта Трихоплакс – очень сильно уплощенное морское животное, обитающее на водорослях. Напоминает амебу, поскольку при движении контуры тела меняются. Движется при помощи жгутиковых клеток, покрывающих тело. Внутри тела располагаются веретеновидные и амебоидные клетки. У трихоплакса обнаружено два способа питания: 1) внеклеточное, 2) фагоцитоз. При первом способе клетки «брюшной» стороны выделяют ферменты, которые переваривают бактерии, покрывающие водоросли. При втором способе пищевые частицы биением жгутиков загоняются на спинную сторону. Здесь эти частицы поглощаются веретеновидными клетками, которые выдвигаются наружу между жгутиковыми клетками. После поглощения пищевой частицы веретеновидная клетка становится амебоидной и погружается внутрь трихоплакса. Размножаются трихоплаксы бесполым и половым способами. Бесполое размножение осуществляется или путем деления тела надвое, или путем отпочковывания «бродяжек». Трихоплакс обладает высокой способностью к регенерации. Трихоплаксы, по-видимому, являются потомками первых многоклеточных животных на Земле. Их строение соответствует строению «фагоцителлы», гипотетического организма, предложенного И.И. Мечниковым для объяснения механизма происхождения многоклеточных животных. Происхождение многоклеточных животных Гипотеза «целялюризации» (Иоринг Хаджи - 1963г.) Многоклеточные животные возникли из одноклеточных полиэнергидных животных путем образования клеточных границ вокруг ядер и прилегающих к ним участков цитоплазмы. Многие протисты, такие как инфузории или слизевики, могут иметь несколько ядер, что подтверждает эту гипотезу. Однако простого наличия нескольких ядер недостаточно для поддержки теории. Множественные ядра инфузорий различны и имеют четко дифференцированные функции. Макроядро служит потребностям организма, а микроядро используется для полового размножения с обменом генетического материала. Синцитиальные слизевики образуются из отдельных амебоидных клеток, подобно синцитиальным тканям некоторых многоклеточных организмов, а не наоборот. Так что, чтобы считаться действительной, эта теория нуждается в наглядном примере и механизме образования многоклеточного организма из ранее существовавшего синцития. Гипотеза симбиотического происхождения Эта теория предполагает, что первые многоклеточные организмы произошли от симбиоза (- форма взаимоотношений организмов, при которой оба организма получают пользу друг от друга) различных видов одноклеточных организмов, каждый из которых играет разные роли. Со временем эти организмы станут настолько зависимыми друг от друга, что не смогут выжить независимо, что в конечном итоге приведет к объединению их геномов в один многоклеточный организм. Каждый соответствующий организм стал бы отдельной линией дифференцированных клеток во вновь создаваемых видах. Однако проблема этой теории заключается в том, что до сих пор неизвестно, как ДНК каждого организма может быть включена в один геном, чтобы составить их как единый вид. Хотя считается, что такой симбиоз имел место, он случался очень редко, и даже тогда геномы эндосимбионтов сохраняли элемент различия, раздельно реплицируя свою ДНК во время митоза видов-хозяев. Например, два или три симбиотических организма, образующих составной лишайник, хотя и зависят друг от друга в отношении выживания, должны воспроизводиться по отдельности, а затем переформироваться, чтобы создать еще один отдельный организм. Гипотезы колониального происхождения Теория предполагает, что симбиоз многих организмов одного и того же вида привел к многоклеточному организму. Преимущество колониальной теории состоит в том, что она была обнаружена независимо в 16 различных типах простейших. Например, во время нехватки пищи амеба группируется в колонии, которая перемещается как единое целое в новое место. Некоторые из этих амеб затем немного отличаются друг от друга. Однако часто бывает трудно отделить колониальных протистов от настоящих многоклеточных организмов, поскольку эти две концепции не различаются; колониальные протисты были названы «плюрицеллюлярными», а не «многоклеточными». Теория гестреи (Геккель – 1872 г.) Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник. Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных. Так развиваются ланцетники и коралловые полипы. Теория фагоцителлы (Мечников – 1878 г.) Фагоцителла не имела рта и кишечника, пищеварение было внутриклеточное. Рот сформировался, как просвет между клетками наружного слоя, ведущий во внутреннюю паренхиму. Располагался он, в отличие от гастреи на заднем конце тела. Кишечника еще не было. Но теперь возникла возможность питаться более крупной добычей: внутренние клетки могли окружать ее, образуя гигантскую пищеварительную вакуоль. Однако для хищничества нужна еще способность ловить добычу. Поэтому хищничать научились только настоящие многоклеточные - после того, как у них возникли мышцы и управляющая ими нервная система. Постепенно у потомков фагоцителлы сформировался постоянный кишечник. По мере увеличения размеров он мог усложняться: возникли боковые карманы, чтобы доставлять пищу к наружным слоям клеток. В дальнейшем у некоторых животных эти карманы могли отделиться, дав начало полости тела - целому. Фагоцителла обитала в толще воды. Нетрудно представить себе, как от нее могли произойти современные группы животных при переходе к жизни на дне. Когда рта еще не было, осевшая на дно фагоцителла "превратилась" в трихоплакса. После появления рта, но до появления кишечника при переходе к ползанию возникли бескишечные турбеллярии. Рот у них сместился на брюхо, и они стали двустороннесимметричными. После появления кишечника часть потомков фагоцителлы перешли к сидячему образу жизни на дне - они превратились в кишечнополостных. Так развиваются обыкновенные губки, гидроидные и сцифоидные стрекающие. Основным кандидатом на роль предка всех многоклеточных является воротничковый жгутиконосец, их клетки снабжены жгутиком, окруженный воротничком. Жгутик создает токи жидкости, позволяющие хоанофлагеллятам плавать в толще воды. Эти же токи пригоняют в воротничковую зону бактерий, которыми хоанофлагелляты питаются. Многие их этих простейших образуют колонии, причем легко переходят из одноклеточного состояния в колониальное и обратно. Теория синзооспоры (Захваткин – 1949 г.) Согласно данной теории многоклеточные также произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются одноклеточные мелкие расселительные стадии - зооспоры. У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия. Отличия от теорий фагоцителлы и гастреи: - Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки. - Сидячий образ жизни примитивных взрослых многоклеточных. - Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения. |