Лекция - происхождение МПП. Механизм развития ПД.. Происхождение мембранного потенциала покоя
 Скачать 306.23 Kb. Скачать 306.23 Kb.
|

|
Лекция 3. ПРОИСХОЖДЕНИЕ МЕМБРАННОГО ПОТЕНЦИАЛА ПОКОЯ При создании мембранного потенциала покоя важную роль играют процессы простой диффузии через белковые каналы в мембране и первично активный транспорт. В основном это диффузия ионов К+ через мембрану клетки из внутриклеточной среды во внеклеточную, что предопределено: 1. Электрохимическим градиентом для K+, Na+, Са2+, Cl-; 2. Избирательно высокой проницаемостью мембраны для К+; 3. Наличием Nа+К+- насоса в мембране. Роль электрохимического градиента для K+, Na+ и Cl- в создании мембранного потенциала на мембране возбудимой клетки Ионы K + , Na+, Cl- проходят через транспортные каналы мембраны согласно электрохимическому градиенту. Этот градиент составляют два компонента: - электрический (статический - в результате того, что мембрана непроницаема для анионов клетки - глутамата, аспартата, органических фосфатов, белков, на внутренней поверхности мембраны образуется избыток отрицательно заряженных частиц, а на наружной – избыток положительно заряженных частиц); - химический градиент концентрации ионов по обе стороны мембраны (концентрация внутри К+ клетки больше, чем вне в 30-50 раз раз, ионов Na+ наоборот, вне клетки больше в 10-20 раз, чем внутри клетки, анионов хлора снаружи больше в 15 раз, а кальция - в 20000 раз). Мембранный потенциал, при котором наблюдается такое равновесие, называется потенциалом равновесия или равновесным потенциалом и рассчитывается согласно уравнению Нернста: Ex=(R·T/z·F) · ln ([X]o/[X]i), где R – газовая постоянная T – температура по Кельвину z – валентность иона F – константа Фарадея ([X]o/[X]I - концентрации ионов снаружи и внутри клетки. Ex – равновесный потенциал. Итак, равновесный потенциал зависит только от концентрации ионов по обе стороны мембраны, но не от свойств ионного канала или механизма проникновения ионов через канал. Избирательно высокая проницаемость мембраны для ионов Ионный ток определяется не абсолютным значением мембранного потенциала, а разницей между мембранным потенциалом и равновесным потенциалом для данного иона. Таким образом, ток через одиночный канал пропорционален разнице между значением мембранного потенциала и значением равновесного потенциала для иона. Проводимость канала прямо пропорциональна току, проходящему через канал и обратно пропорциональна потенциалу. Проницаемость канала определяется особенностями прохождения ионов через канал. Транспортные белки обладают высокой избирательной проницаемостью для ионов, особенно для К+, Na+, Cl- . Было рассчитано, что в изолированном гигантском аксоне кальмара соотношение проницаемостей РK+ : РNa+ :Р Cl- относительно калия для некоторых ионов можно представить как 1 : 0,04 : 0, 45. По строению все ионные каналы похожи друг на друга, состоят из разного количества субъединиц, каждая из которых состоит из повторяющихся доменов, которые в свою очередь состоят из шести спиральных сегментов, пронизывающих мембрану. Эта структура сворачивается с образованием поры канала за счет наружных аминокислотных петель между 5 и 6 сегментами. Сенсор располагается в четвертом сегменте. Возможно, что все каналы произошли от одного источника – калиевого или кальциевого канала простейших. Различают каналы неуправляемые – безворотные или каналы утечки и управляемые – одноворотные или двуворотные. Управляемые каналы могут быть потенциалчувствительными, лигандчувствительными или механочувствительными. Ионные потенциалзависимые каналы - это ионные каналы, которые открываются и закрываются в ответ на изменение мембранного потенциала, например, натриевые каналы, кальциевые каналы. Лигандзависимые каналы - ионные каналы, расположенные в постсинаптической мембране в нервно-мышечных соединениях. Механозависимые каналы. Способность клеток трансформировать механическое воздействие в электрический и химический сигналы (механотрансдукция) обеспечивает восприятие звука и вибрации, гравитации, ускорения, скорости, давления, прикосновения, изменения формы и объема клетки, ее местоположения относительно внеклеточного матрикса и окружающих клеток. Таким образом, избирательность ионных каналов обусловлена: - Геометрией каналов (размер устья, селективного фильтра, отрицательный заряд на поверхности канала); - Воротным механизмом каналов; - Электрическим зарядом окружающих структур ( потенциалчувствительные каналы) или лигандом, т.е. либо внутриклеточным метаболитом, либо внутриклеточным вторичным мессенджером либо внеклеточным лигандом, имеющим сродство к определенным рецепторам на мембране ( хемочувствительные или лигандзависимые каналы), структурным изменением мембраны при растяжении ( механочувствительные каналы). Роль Nа+К+- насоса в мембране при создании мембранного потенциала покоя возбудимых клеток Неравенство концентраций ионов К+ и Na+ внутри и снаружи клетки ( волокна) поддерживается специальным механизмом (т. Н. натриевым насосом), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ. Работа таких механизмов обеспечивается, как правило, энергией, выделяемой при расщеплении аденозинтрифосфорной кислоты (АТФ). Активный перенос Na+ из клетки сопряжён с транспортом К+ в обратном направлении и осуществляется особой ферментной системой – транспортной Na, К, - стимулируемой аденозинтрифосфатазой, локализованной в клеточной мембране. Последняя, гидролизуя АТФ, высвобождает энергию, которая и затрачивается на активный перенос катионов. Работа насоса в целом зависит от уровня метаболизма клетки. Nа+,К+- насос переносит из клетки ионы 3 Nа+, а вносит 2 К+ против градиента концентрации. В связи с этим перенос веществ через мембрану сопровождается изменением мембранного потенциала в среднем на 10 мВ, поэтому такой вид транспорта является электрогенным. Переносчик теряет сродство к Nа+, но приобретает сродство к К+. В результате меняется конформация белка и К+ пассивно вносится внутрь клетки. МЕХАНИЗМЫ РАЗВИТИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ Потенциал действия и фазы его развития Исходя из вышесказанного, клеточная мембрана возбудимых систем мембрана несет заряд, величина которого колеблется от – 60 – 90 мВ. Уменьшение заряда мембраны называется деполяризацией, а увеличение заряда – гиперполяризацией При нанесении раздражения с силой, равной некоторой критической величине, достигается критический уровень деполяризации (Екр или КУД) и развивается быстрая деполяризация или ПД, или спайк. Разница между Ео и Екр. называется порогом деполяризации или пороговым потенциалом (ΔЕ). Чем меньше ΔЕ, тем выше возбудимость. Порог деполяризации – одна из характеристик мембраны, отражает ее особенности и функциональное состояние и является мерой возбудимости. При изменении функционального состояния мембраны наблюдаются разные ΔЕ. При сдвиге МПП до критического уровня мембрана возбудимых структур резко изменяет свою ионную проницаемость. Под влиянием трансмембранных ионных потоков внутренняя поверхность мембраны не только теряет свой отрицательный потенциал, но и приобретает положительный заряд в момент пика, достигающий +30; +50 мВ (овершут), т.е. происходит инверсия потенциала мембраны. При достижении критического уровня деполяризации открываются все имеющиеся натриевые каналы по закону " все или ничего", поэтому при сверхпороговом раздражении критический уровень деполяризации достигается быстрее, однако амплитуда ПД остается такой же. Возникающий при этом потенциал получил название потенциала действия ( его величина в среднем 110 мВ). 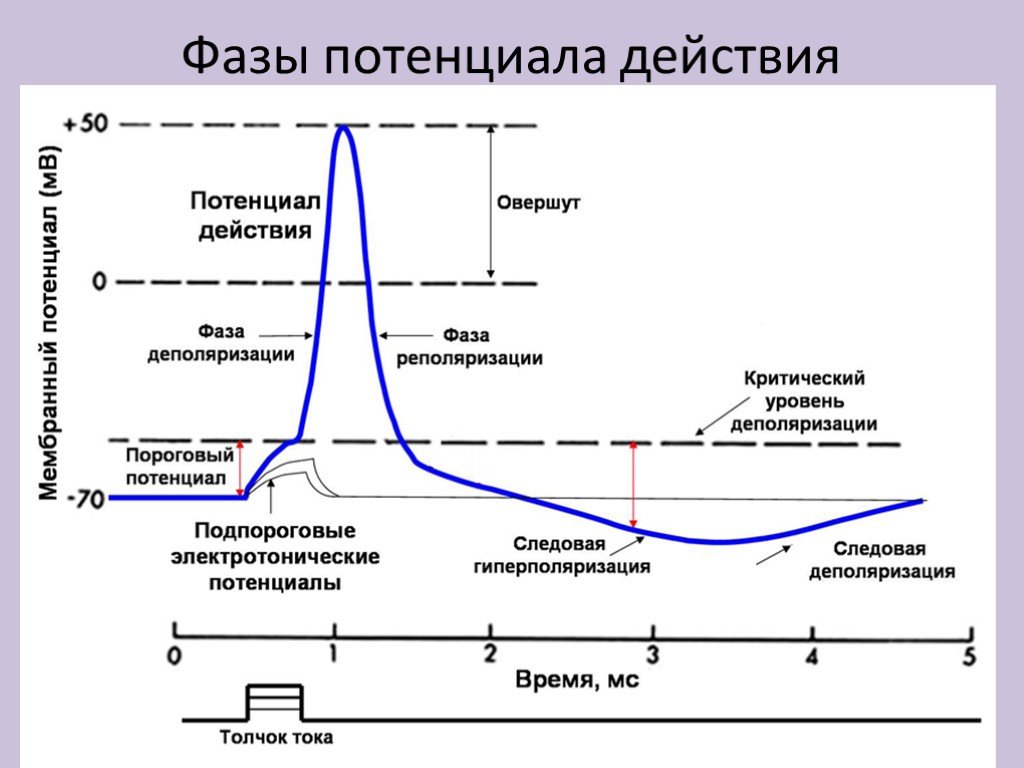 Таким образом, потенциал действия – это быстрое колебание МПП, возникающее при возбуждении нервных, мышечных и секреторных клеток. В развитии потенциала действия прослеживается несколько основных фаз: 1.Фаза быстрой деполяризации 2.Фаза реполяризации 3.Фаза следовой деполяризации или отрицательный следовой потенциал (замедление реполяризации) 4.Фаза следовой гиперполяризации или положительный следовой потенциал. Фаза быстрой деполяризации. В соответствии с положениями мембранной теории, возникновение потенциала действия объясняется тем, что при достижении деполяризации критического уровня в мембране открываются все натриевые каналы. Изменение электрического поля мембраны приводит к открыванию воротных механизмов - активационных ворот (m) в потенциалзависимом натриевом канале (рис. 32). При этом проницаемость для Na+ резко (в 20 раз) увеличивается по сравнению с проницаемостью для К+, и в 500 раз по сравнению с исходной проницаемостью Na+ в покое (рис. 31Б). Проницаемость для К+ практически остается на прежнем уровне (K+ : Na+ : Cl- как 1:20:0,45). Ионы Na+ по концентрационному и электрохимическому градиенту лавинообразно устремляются внутрь возбудимых клеток, неся с собой положительные заряды. Однако фаза быстрой деполяризации кратковременна (например, в нервной клетке она составляет всего 0,5 мс – 1мс). Фаза реполяризации. Пик ПД (+30 мВ) обусловлен максимальным повышением проницаемости для ионов К+ (с началом деполяризации открываются дополнительно медленные одноворотные калиевые каналы) и резким снижением проницаемости для Na+ (закрываются медленные инактивационные ворота h в натриевом канале). Наступает более длительная фаза реполяризации, обусловленная Na+-инактивацией и повышением проницаемости для К+. Фаза следовых потенциалов. В конечной фазе возбуждения происходит замедление восстановления мембранного потенциала покоя, и при этом регистрируются следовые реакции в виде следовой деполяризации и гиперполяризации, обусловленные медленным восстановлением исходной проницаемости для ионов К+. Итак, ПД развивается в основном в результате пассивного транспорта ионов Na+ внутрь клетки согласно электрохимическому градиенту. Для поддержания концентрационного градиента после серии разрядов включается активная деятельность мембранных АТФаз (Na+,К+-насосов), направленных на восстановление измененного концентрационного градиента Na+ и К+. Условия возникновения возбуждения в возбудимых системах Не всякое раздражение приводит к возникновению возбуждения, т.е. развитию потенциала действия. Необходимы определенные условия для возникновения возбуждения в возбудимых системах: 1). Структурно-функциональное состояние мембраны ( достаточный уровень МПП, определенный уровень Екр. или порогового потенциала (ΔЕ), уровень К+, Na+проницаемости) 2). Значение параметров раздражителя ( сила, время, градиент нарастания силы во времени), достаточное для возникновения возбуждения. Минимальная сила раздражителя, способная вызвать возбуждение, называется порогом раздражения. В связи с данным определением можно выделить подпороговое, пороговое и сверхпороговое раздражение. При нанесении раздражения с силой <50% пороговой величины происходит пассивная деполяризация мембраны, называемая электротоническим ответом или электротоном. При нанесении раздражения с силой 50%- 99,9% пороговой величины к пассивному изменению электрического поля мембраны добавляется активное изменение в форме повышения натриевой проводимости. Часть Na+каналов открывается. Активные подпороговые изменения МПП называются локальным ответом. Локальные ответы: • не способны к распространению, • градуальны (чем больше раздражение, тем больше ответ). Пример: ВПСП (возбуждающий постсинаптический потенциал), ТПСП ( тормозной постсинаптический потенциал), рецепторный потенциал. Итак, мерой возбудимости можно считать порог раздражения, хотя этот показатель возбудимости ткани относится к характеристике раздражителя, а не возбудимой системы. При пародонтозе, например, порог раздражения зуба ниже нормы, что свидетельствует о повышении возбудимости. При пульпитах, наоборот, отмечается повышение порога раздражения. Максимальное снижение возбудимости наблюдается при гибели пульпы. Но для характеристики возбудимости тканей важно учитывать не только пороговую силу раздражителя, но и время действия раздражителя на ткань. Существует определенная зависимость между временем действия раздражителя и его силой. Эта зависимость для электрического тока в графическом выражении ( гипербола) получила название кривой «сила-длительность». По имени ее авторов – кривая Гоорвейга-Вейса-Лапика (1892, 1901, 1909). Минимальная величина силы раздражителя, вызывающая возбуждение, называется абсолютным порогом силы, или реобазой (от греч. rheos - течение, поток и basis - ход, движение; основание). С другой стороны, раздражитель должен действовать не меньше определенного времени. Уменьшение времени действия раздражителя ниже критического значения приводит к тому, что раздражитель любой интенсивности не оказывает эффекта (высокочастотный переменный ток >10 кГц дает только тепловой эффект при коротком времени действия). Минимальная величина времени действия раздражителя, вызывающая возбуждение, называется абсолютным порогом времени. С учетом действия двух параметров раздражителя для характеристики возбудимости ткани ввели понятие полезного времени. Полезное время - это минимальное время, в течение которого должен действовать раздражитель пороговой силы с тем, чтобы вызвать возбуждение. Приближение кривой асимптотически к линии, параллельной абсциссе, не позволяет достаточно точно определять полезное время, т.к. незначительные отклонения реобазы, отражающие изменения функционального состояния биологических мембран в покое, сопровождаются значительными колебаниями времени раздражения. В связи с этим Лапик предложил измерять другую условную величину - хронаксию (от греч. chronos - время и axia - цена, мера). Хронаксия – время (отрезок АЕ), в течение которого должен действовать раздражитель удвоенной реобазы (отрезок АF), чтобы вызвать возбуждение. Использование этого критерия позволяет точно измерить временные характеристики возбудимых структур, поскольку измерение происходит на крутом изгибе гиперболы. Чем меньше хронаксия, тем больше возбудимость. Например, хронаксия нервных волокон ниже, чем мышечных. Хронаксиметрия используется при оценке функционального состояния нервно-мышечной системы у человека ( в случае повреждения нерва и его перерождения определяют истинную хронаксию мышцы, которая намного превышает таковую до травмы), в частности челюстно-лицевой области. Показатели хронаксии и реобазы могут значительно меняться при невритах и невралгиях тройничного и лицевого нервов, миозитах мимической и жевательной мускулатуры. Таким образом, количественная оценка физиологических свойств возбудимых биосистем в клинической практике производится опосредованно по характеристикам раздражителя. Третьим параметром раздражителя является градиент нарастания силы во времени.  Факторы, определяющие характер ответа возбудимых биосистем на раздражение Рассмотрим факторы, определяющие характер ответа возбудимых биосистем на раздражение. Первым фактором является влияние параметров раздражителя ( силы, времени действия, крутизны нарастания силы во времени). Вторым фактором, определяющим характер ответа возбудимых биосистем, является функциональное состояние биосистемы. Рассмотрим влияние параметров раздражителя. Характер влияния параметров раздражителя описывается тремя законами раздражения: • законом силы; • законом времени; • законом градиента нарастания силы во времени. Для возникновения возбуждения должны быть достаточными сила, время действия раздражителя, крутизна нарастания силы раздражителя во времени. При подпороговом раздражении не возникает возбуждения, так как мы уже знаем о механизме формировании локального ответа на подпороговый раздражитель. При пороговом раздражении – возникает возбуждение, развивается ПД, обусловленный открытием сразу всех натриевых каналов, поэтому действие сверхпороговых раздражителей не вызовет увеличения амплитуды ответа. Таким образом, для клетки закон силы и времени действует по принципу «все или ничего». Перейдем к законам раздражения для ткани. Ответ ткани на раздражитель, прежде всего, зависит от различной возбудимости клеток ткани. Для ткани законы силы и времени действуют по принципу «силовых отношений». При подпороговом раздражении, например, мышца не отвечает сокращением. При пороговом стимуле – ответ ткани, т.е. самые возбудимые клетки пришли в возбуждение и сократились (здесь порог для мышцы и для клетки). При повышении стимула ответят самые возбудимые клетки по закону «все или ничего» и ответят другие клетки, для которых порог более высокий, т.е. увеличится количество сокращающихся клеток – возрастет суммарный ответ мышцы. В конце концов, возникнет ситуация, когда все элементы мышцы отреагируют. Дальнейшего увеличения амплитуды ответа не произойдет. В этом случае различают: 1). Подпороговые раздражители 2). Пороговые раздражители 3). Сверхпороговые раздражители – субмаксимальные, максимальный, супермаксимальные раздражители. Та же закономерность существует и для градиента нарастания силы во времени действия раздражителя с той разницей, что беспредельно нарастать крутизну нельзя, так как дальше идет прямоугольный импульс. Вторым фактором, определяющим характер ответа возбудимых биосистем, является функциональное состояние биосистемы. Оказывается, что даже в процессе возбуждения изменяется возбудимость мембраны, что проявляется как физиологический компонент возбуждения. 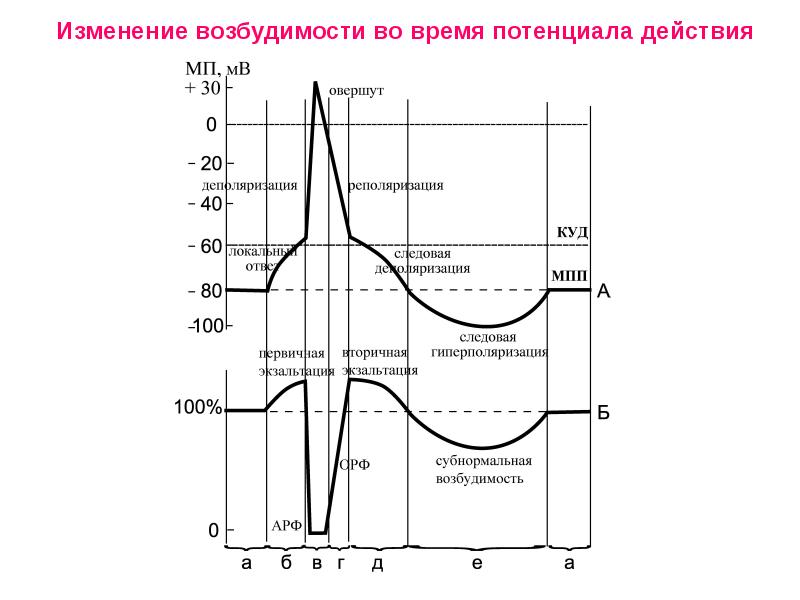 Изменение возбудимости во время возбуждения имеет фазный характер, в связи с чем выделяют несколько фаз: 1. Фаза абсолютной рефрактерности 2. Фаза относительной рефрактерности 3. Фаза супернормальной возбудимости (экзальтации) 4. Фаза субнормальной возбудимости Фаза абсолютной рефрактерности - это полная невозбудимость клетки. Соответствует пику ПД и продолжается в клетках 1-2 и более мс. В этот период активационные m-ворота натриевых каналов открыты по закону " все или ничего", поэтому дополнительное раздражение не может вызвать еще большее открывание каналов и появление новых ПД. Фаза относительной рефрактерности – это период восстановления возбудимости, когда сильное раздражение может вызвать новое возбуждение. Пониженная возбудимость является следствием все еще повышенной проницаемости для К+ и избыточного выхода его из клетки, препятствующего деполяризации клетки при действии раздражителя, и все еще большим количеством инактивированных натриевых каналов. Только сильное раздражение может вызвать возбуждение. Фаза экзальтации – это период повышенной возбудимости. Он соответствует отрицательной следовой деполяризации. Очередной ПД можно вызвать более слабым раздражителем, т.к. мембранный потенциал несколько меньше обычного и оказывается ближе к критическому уровню деполяризации. Фаза субнормальной возбудимости – это период пониженной возбудимости, Соответствует периоду следовой гиперполяризации, когда мембранный потенциал больше обычного и дальше отстоит от критического уровня деполяризации. Лабильность как одно из общих свойств возбудимых систем Впервые понятие лабильности как функциональной подвижности возбудимых тканей ввел русский физиолог Н.Е. Введенский. На современном этапе развития физиологии лабильность рассматривается как способность биосистемы в течение времени развертывать одиночный процесс возбуждения. Мерой лабильности является максимальное число возбуждений или ПД, которое способна генерировать возбудимая биосистема за единицу времени в связи с навязанным ритмом возбуждения. Лабильность нервного волокна – 1000 имп/с, мышцы – 200 имп/с. Лабильность возбудимых биосистем практически обусловлена длительностью фазы абсолютной рефрактерности. Так, фаза абсолютной рефрактерности нервного волокна равна в среднем 1 мс, то есть в 1 секунду при ритмическом раздражении с частотой 1000 Гц нерв может воспроизвести 1000 импульсов. Фаза абсолютной рефрактерности у мышцы – 5 мс, то есть в 1 секунду мышца может воспроизвести 200 возбуждений. Лабильность также является мерой возбудимости. Протекание процесса возбуждения во времени характеризует в возбудимых тканях и лабильность и хронаксия. Какой их этих показателей дает более полную характеристику процесса возбуждения? Хронаксия – это время, в течение которого должен действовать ток, силой в 2 реобазы, чтобы вызвать возбуждение. В таком случае, хронаксия характеризует только начальную стадию – возникновение импульса возбуждения, а лабильность – протекание всего импульса. Кроме того, хронаксия связана с одиночным возбуждением, а лабильность – с множестваом импульсов возбуждения, взаимодействующих друг с другом. Поэтому лабильность более полно характеризует протекание возбуждения во времени. Итак, общими свойствами возбудимых биосистем являются: 1. Возбудимость 2. Лабильность 3. Проводимость При этом мерой возбудимости могут служить: • Порог раздражения (характеристика раздражителя) • Хронаксия (характеристика раздражителя) • Пороговый потенциал (характеристика мембраны) • Лабильность (характеристика мембраны) |