Ботаника. ботаника. Рефераты по учебной дисциплине Ботаника
 Скачать 2.82 Mb. Скачать 2.82 Mb.
|

|
РЕФЕРАТЫ по учебной дисциплине «Ботаника» 2021г. Содержание:
1 (15). Опишите образование, строение и функции пор. Приведите рисунки. Что такое плазмодесмы? Во вторичной стенке имеется большое количество пор. Каждая пора представляет собой канал в том месте клеточной оболочки, в котором над первичным поровым полем не откладывается вторичная оболочка. Первичное поровое поле — это небольшой участок тонких смежных стенок двух клеток, состоящий из первичной оболочки и клеточной пластинки, пронизанный плазмодесмами. Поры возникают парно в смежных клетках соседних клеток и разделены замыкающей трёхслойной плёнкой (поровой мембраной). Различают поры: 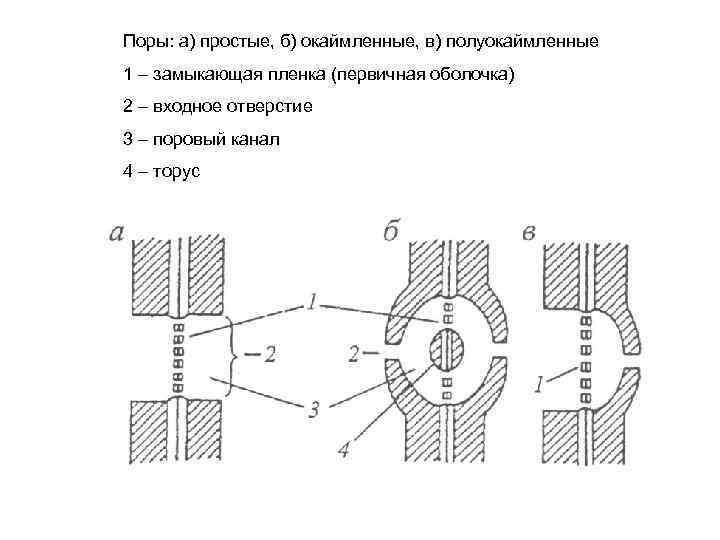 Простые поры представляют собой каналы во вторичной оболочке паренхимных клеток и склереид, имеющие одинаковую ширину на всем протяжении. Простые поры представляют собой каналы во вторичной оболочке паренхимных клеток и склереид, имеющие одинаковую ширину на всем протяжении.Окаймлённые поры — это поры, окаймление которых составляет куполообразно приподнятая над поровой мембраной вторичная оболочка. В плане такая пора имеет вид двух окружностей, наружная из которых соответствует окаймлению, а внутренняя — отверстию, открывающемуся в полость клетки. Характерны для водопроводящих элементов, представленных мёртвыми клетками. Полуокаймлённые поры — пара пор, одна из которых — простая, другая — окаймлённая. Образуется в смежных стенках трахеид хвойных и паренхимных клеток древесинных лучей. Слепые поры представляют собой каналы во вторичной оболочке только одной из двух соседних клеток, такие поры не функционируют. Ветвистые поры — поры, разветвлённые на одном из концов вследствие слияния двух или нескольких простых пор в процессе утолщения вторичной оболочки. Щелевидные поры — поры с отверстиями в виде косой щели; образуются в клетках прозенхимы, например, волокнах древесины. Обмен веществами между ядром и цитоплазмой клетки осуществляется посредством ядерных пор — транспортных каналов, пронизывающих двухслойную ядерную оболочку. Переход молекул из ядра в цитоплазму и в обратном направлении называется ядерно-цитоплазматическим транспортом. Ядерно-цитоплазматическим транспортом называется материальный обмен между клеточным ядром и цитоплазмой клетки. Ядерно-цитоплазматический транспорт можно разделить на две категории: активный транспорт, требующий затрат энергии, а также специальных белков-рецепторов, и пассивный транспорт, протекающий путем простой диффузии молекул через канал ядерной поры. [1]  Плазмодесмы — цитоплазматические мостики, соединяющие соседние клетки растений. Плазмодесмы проходят через канальцы поровых полей первичной клеточной стенки. Благодаря плазмодесмам растительные клетки образуют многоклеточные структуры — симпласты, в пределах которых между клетками напрямую передаются ионы и малые молекулы (в том числе сигнальные молекулы). Плазмодесмы могут закрываться и открываться. Многие вирусы растений увеличивают размер пор плазмодесм, чтобы обеспечить транспорт вирусных частиц через клетки. Плазмодесмы — цитоплазматические мостики, соединяющие соседние клетки растений. Плазмодесмы проходят через канальцы поровых полей первичной клеточной стенки. Благодаря плазмодесмам растительные клетки образуют многоклеточные структуры — симпласты, в пределах которых между клетками напрямую передаются ионы и малые молекулы (в том числе сигнальные молекулы). Плазмодесмы могут закрываться и открываться. Многие вирусы растений увеличивают размер пор плазмодесм, чтобы обеспечить транспорт вирусных частиц через клетки.Плазмодесмы образуются в ходе последнего этапа деления растительных клеток — цитокинезе, когда дочерние клетки окончательно отделяются друг от друга. В новообразованных клеточных стенках остаются поры, соединяющие цитоплазмы соседних клеток. Плазмодесмы формируются, если при формировании клеточной пластинки в неё попадают трубочки эндоплазматического ретикулума (ЭПР). При этом мембраны соседних клеток и трубочки ЭПР не сливаются друг с другом. Вместо этого трубочка ЭПР оказывается разделённой на две части, которые находятся в соседних клетках и соединены узким перешейком. Перешеек же залегает в канале в клеточных стенках соседних клеток и окружён цитоплазмой. Его также называют десмотубулой. Плазмодесмы, сформированные при цитокинезе, называются первичными. Однако иногда плазмодесмы соединяют клетки, не являющиеся продуктом одного деления; такие плазмодесмы называются вторичными. Вторичные плазмодесмы образуются при локальном утоньшении стенки между двумя клетками, и в образующееся отверстие каким-то образом вставляется трубочка ЭПР. Типичная растительная клетка имеет от 103 до 105 плазмодесм. В меристемах растений плазмодесмы обычно расположены диффузно. В клетках корневой меристемы лука на 1 мкм² приходится 6—7 плазмодесм, а у мужского папоротника — 10—20 плазмодесм. Плазмодесмы объединяют растительные клетки в одну большую структуру, которая называется симпласт. В пределах симпласта через плазмодесмы проходят электрические сигналы, ионы и небольшие водорастворимые молекулы, включая аминокислоты, сахара, мРНК, малые интерферирующие РНК и белки (в том числе транскрипционные факторы). Ионы и молекулы проходят через плазмодесмы с помощью простой диффузии. Диаметр пор плазмодесм может изменяться, обеспечивая возможность настройки избирательной проницаемости плазмодесм. Иногда молекулы доставляются к плазмодесмам с участием актиновых филаментов: плазмодесмы, как правило, располагаются в тех местах клеточной стенки, где сосредоточен актиновый цитоскелет. Актиновые филаменты служат основой очень активной системы, обеспечивающей перенос метаболитов на значительные расстояния по цитоплазме вакуолизированных растительных клеток. Нити актина направляют ток цитоплазмы, который и направляет молекулы к плазмодесмам. Актиновые нити могут прикрепляться к клеточным стенкам при помощи особой, уникальной для растений формы миозина. В порах плазмодесм тоже обнаружены тонкие актиновые нити и молекулы миозина. Возможно, диаметр пор меняется именно благодаря их сокращению и расширению. Многие патогены растений — вироиды и вирусы — переносятся между клетками посредством плазмодесм, несмотря на свои большие размеры. Дело в том, что вирусы экспрессируют белки, которые расширяют поры плазмодесм, делая возможным транспорт даже довольно крупных вирусных частиц. Механизм действия этих белков пока не известен. [2] 2 (61). Переход ко вторичному строению корня. Приведите рисунок. Вторичное анатомическое строение - результат работы вторичных латеральных меристем: камбия и феллогена. Переход от первичного анатомического строения к вторичному осуществляется в зоне проведения. При переходе к вторичному анатомическому строению происходит формирование вторичной флоэмы, вторичной ксилемы и радиальных (сердцевинных) лучей. Вторичная ксилема и флоэма, также, как и радиальные паренхимные лучи, формируются в результате деятельности камбиального кольца. Камбиальное кольцо закладывается в центральном цилиндре. Вначале внутрь от первичной флоэмы начинается деление паренхимных клеток остаточного прокамбия. Возникшие разобщённые дуги делящихся клеток смыкаются с клетками перицикла, в который они упираются, и клетки которого также начинает делиться. Возникает извилистое кольцо камбия. Участки камбиального кольца, имеющие различное происхождение, функционируют различно. Камбий, возникший из прокамбия между первичными флоэмой и ксилемой, образует вторичные флоэму и ксилему. Камбий перициклического происхождения даёт паренхиму радиальных лучей. Образование вторичных тканей в центральном цилиндре корня ведёт к увеличению его объема и, естественно, влечёт за собой разрушение первичной коры. В это время из клеток перицикла формируется феллоген. Он образует перидерму, которая отделяет первичную кору от центрального цилиндра. Первичная кора отмирает и слущивается. Этот процесс называется "линька корня". На поверхности корня остаётся перидерма. Таким образом, переход ко вторичному анатомическому строению корня связан с: 1 - сбрасыванием первичной коры ("линька корня"); 2 - заложением камбия и образованием вторичной ксилемы, вторичной флоэмы и радиальных паренхимных лучей; 3 - образованием феллогена из перицикла с последующем формированием перидермы. Между перидермой и камбием располагается флоэма - вторичная кора, под камбием - ксилема, или древесина. Порядок работы: Постоянный препарат поперечного среза корня вторичного строения тыквы обыкновенной изучают при малом увеличении. Находят на срезе чётко выделяющиеся четыре участка вторичной ксилемы. Стенки клеток ксилемы - одревесневшие, на препарате они окрашены в красный цвет. Крупные округлые просветы – полость сосудов. Между участками вторичной ксилемы располагаются радиальные лучи, состоящие из паренхимных клеток с целлюлозными стенками. Лучи сужаются по направления к центру корня и упираются в вершины лучей первичной ксилемы, клетки которой, также, как и вторичной ксилемы, одревесневшие. Четыре луча первичной ксилемы (ксилема тетрархная) по направлению к центру корня слегка расширяются и упираются в крупный центральный сосуд. Находят зону камбия. Она примыкает снаружи к участкам вторичной ксилемы. Клетки камбиальной зоны располагаются в несколько рядов. Рассматривают расположенную снаружи от камбия кору корня. Здесь, ближе к камбию, видна вторичная флоэма, дальше от неё - первичная флоэма. Границу между ними различить практически невозможно. Все клетки флоэмы имеют целлюлозные стенки и окрашены в голубой цвет. Участков флоэмы, как и вторичной ксилемы, четыре. Между ними расположены паренхимные клетки радиальных лучей. Переводят микроскоп на большое увеличение и рассматривают строение камбиальной зоны. Обращают внимание на взаимное расположение клеток, густую цитоплазму и крупные ядра. Рассматривают вторичную покровную ткань - пробку, входящую в состав перидермы. Клетки пробки имеют светло-коричневую окраску. [3] 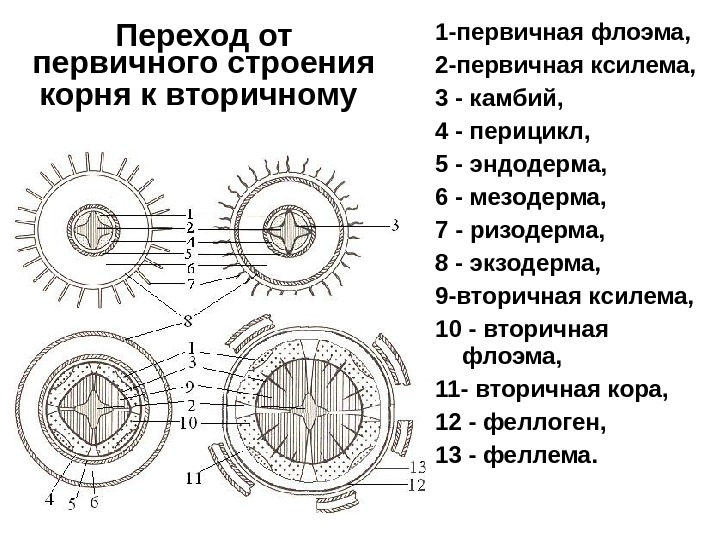 3. (96). Самоопыление. Клейстогамия, дихогамия, гетеростилия. Приведите примеры ирисункиия. Самоопыление — форма гомогамии, тип опыления у высших растений. При самоопылении пыльца из пыльников переносится на рыльце пестика того же самого цветка или между цветками одного растения. К самоопыляемым растениям относятся горох, фиалки, пшеница, помидоры, ячмень, фасоль, нектарин. Различают несколько способов самоопыления: автогамия — перенос пыльцы на рыльце пестика того же хазмогамного (открытого) цветка. Реализуется разными способами: в бутонах (бутонная автогамия), при прямом соприкосновении рыльца и пыльников (контактная автогамия), при высыпании пыльцы из пыльников и оседании ее на рыльце под действием собственной тяжести (гравитационная автогамия), ветра (ветроавтогамия), мельчайших насекомых, обитающих в цветке (трипс-автогамия). Бутонная автогамия встречается реже, чем другие способы автогамии, отмечена у представителей семейств гвоздичные, бобовые, розовые, норичниковые, злаки. Контактную автогамию можно наблюдать у седмичника европейского, цирцеи альпийской, майника двулистного. Обязательная контактная автогамия наблюдается у копытня европейского. Гравитационная автогамия наблюдается у одноцветки. Гравитационной автогамии может способствовать ветер, поскольку при раскачивании и сотрясении растений пыльца высыпается через отверстия (поры), имеющиеся в пыльниках вересковых и грушанковых. Трипс-автогамия встречается у разных растений, чаще всего у сложноцветных с мелкими трубчатыми цветками. гейтоногамия — опыление происходит в пределах одного растения. Может совершаться различными путями. Например, под влиянием собственной тяжести пыльца с верхних цветков соцветия может попадать на рыльца нижерасположенных цветков. Таким образом происходит опыление у многих вересковых. Гейтоногамии может способствовать ветер или животные, посещающие цветки. Гейтоногамия возможна также при непосредственном контакте восприимчивых рылец одних цветков со вскрывшимися пыльниками соседних цветков. Подобный контакт отмечается в густых соцветиях, состоящих из множества мелких цветков, находящихся в разной стадии цветения (некоторые зонтичные, астровые, капустные, гвоздичные, маревые). Клейстогамия — опыление осуществляется в никогда не открывающихся цветках. Это явление широко распространено у дикорастущих и культурных растений. Обнаружено у почти 700 видов растений из почти 60 семейств. Клейстогамия наблюдается у арахиса, некоторых видов фиалок, гусмании, кислицы обыкновенной, кактусов рода фрайлея.  Самоопыление (в любой форме) рассматривается как вторичное явление, вызванное крайними условиями среды, неблагоприятными для перекрестного опыления. В таких случаях оно выполняет страхующую функцию, выступая в качестве резервного способа опыления. Чаще всего случаи самоопыления отмечаются у растений тундр, темнохвойной тайги, высокогорий, пустынь и у раноцветущих весенних растений. [4] Самоопыление (в любой форме) рассматривается как вторичное явление, вызванное крайними условиями среды, неблагоприятными для перекрестного опыления. В таких случаях оно выполняет страхующую функцию, выступая в качестве резервного способа опыления. Чаще всего случаи самоопыления отмечаются у растений тундр, темнохвойной тайги, высокогорий, пустынь и у раноцветущих весенних растений. [4]Клейстогамия — способ самоопыления, при котором опыление происходит в закрытых, никогда не открывающихся цветках, тем не менее, завязываются плоды и семена. Больше всего клейстогамных видов в семействах: злаки, фиалковые, бобовые, орхидные, акантовые, коммелиновые, никтагиновые, кактусовые. Самым крупным родом клейстогамных растений является фиалка, много клейстогамных растений отмечено также в роде ковыль и ряде других родов семейства злаки, из других семейств растений - у рода подорожник (Plantago), леспедеца, фрайлея. Клейстогамия чаще наблюдается у однолетников, чем у многолетников. 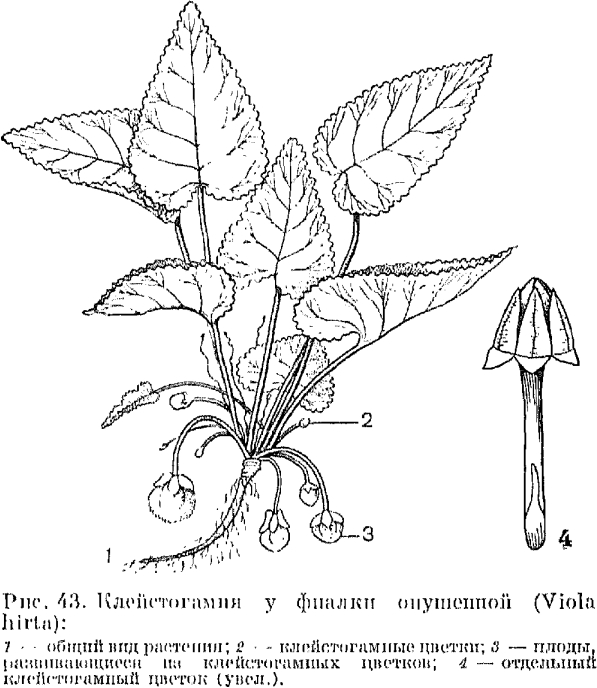 Клейстогамия имеет экологический характер и вызывается неблагоприятными факторами среды. Эти факторы многочисленны и часто противоречивы: клейстогамию могут вызывать недостаток и избыток воды в окружающей среде, повышенная или пониженная относительная влажность воздуха, затенение, низкая или высокая температура, бедность почвы, почвенная засуха, несоответствие длины фотопериода и т.д. Правильное объяснение действия многих факторов затрудняется тем, что на одном и том же растении хазмогамные цветки (раскрывающиеся цветки) появляются вслед за клейстогамными и наоборот. У растений сначала формируются клейстогамные цветки, а потом хазмогамные. У фиалки, опушенной хазмогамные цветки образуются ранней весной, они имеют нормально развитые тычинки и завязи, но, как правило, остаются бесплодными, а после их увядания развиваются крошечные клейстогамные цветки, сидящие на коротких цветоножках. У некоторых растений одновременно развиваются и хазмогамные, и клейстогамные цветки. [5] Клейстогамия имеет экологический характер и вызывается неблагоприятными факторами среды. Эти факторы многочисленны и часто противоречивы: клейстогамию могут вызывать недостаток и избыток воды в окружающей среде, повышенная или пониженная относительная влажность воздуха, затенение, низкая или высокая температура, бедность почвы, почвенная засуха, несоответствие длины фотопериода и т.д. Правильное объяснение действия многих факторов затрудняется тем, что на одном и том же растении хазмогамные цветки (раскрывающиеся цветки) появляются вслед за клейстогамными и наоборот. У растений сначала формируются клейстогамные цветки, а потом хазмогамные. У фиалки, опушенной хазмогамные цветки образуются ранней весной, они имеют нормально развитые тычинки и завязи, но, как правило, остаются бесплодными, а после их увядания развиваются крошечные клейстогамные цветки, сидящие на коротких цветоножках. У некоторых растений одновременно развиваются и хазмогамные, и клейстогамные цветки. [5]Дихогамия — явление неодновременного созревания женских и мужских органов в цветке покрытосеменных растений. Дихогамия проявляется в двух формах: протандрии (протерандрии) — более раннего созревания пыльников с пыльцой и протогинии — более раннего созревания пестиков. Дихогамия является одним из приспособлений к перекрестному опылению и препятствием к самоопылению. Более распространенной формой дихогамии является протандрия (злаки, гвоздичные, гераниевые, мальвовые и многие др.), в то время как протогиния описана только у некоторых представителей розоцветных, зонтичных, крестоцветных. Гетеростилия - это уникальная форма полиморфизма и геркогамии у цветов. У гетеростильных видов в популяции существуют два или три морфологических типа цветков, называемых «морфами». На каждом отдельном растении все цветы имеют одинаковую форму. Цветочные морфы различаются длиной пестика и тычинок, и эти признаки не являются непрерывными. Фенотип морфа генетически связан с генами, ответственными за уникальную систему самонесовместимости, называемую гетероморфной самонесовместимость, то есть пыльца цветка одной морфы не может оплодотворять другой цветок той же морфы.  Длина тычинок и пестиков у гетеростильных цветков адаптирована для опыления разными опылителями или разными частями тела одного и того же опылителя. Таким образом, пыльца, происходящая из длинной тычинки, будет достигать в основном длинных, а не коротких пестиков, и наоборот. Когда пыльца переносится между двумя цветками одной и той же морфы, оплодотворение не происходит из-за механизма самонесовместимости, если только такой механизм не нарушается факторами окружающей среды, такими как возраст цветка или температура. Длина тычинок и пестиков у гетеростильных цветков адаптирована для опыления разными опылителями или разными частями тела одного и того же опылителя. Таким образом, пыльца, происходящая из длинной тычинки, будет достигать в основном длинных, а не коротких пестиков, и наоборот. Когда пыльца переносится между двумя цветками одной и той же морфы, оплодотворение не происходит из-за механизма самонесовместимости, если только такой механизм не нарушается факторами окружающей среды, такими как возраст цветка или температура.Гетеростилия у первоцвета обыкновенного (Primula vulgaris). 1 — тычинки, 2 — столбики с рыльцами на конце. Слева — длинностолбчатый цветок, справа — короткостолбчатый. [6] 4 (131). Практическое значение грибов сапрофитов в природе и жизни человека. Микориза. |