2 коллоквиум. Роль воды в жизни растений. Молекулярная структура и физические свойства воды
 Скачать 73.01 Kb. Скачать 73.01 Kb.
|

|
2 коллоквиум. Водный обмен растений
Для нормальной физиологической деят-ти клетка растения должна быть полностью насыщена водой (или близка к насыщению). В жизни клетки вода имеет значение для поддержания структуры цитоплазмы путем гидратации ее коллоидов и тургорного состояния. Кроме этого, вода необходима как среда для протекания обмена веществ, так как биохимические реакции возможны только между веществами, находящимися в растворенном состоянии. Вода является средой для переноса веществ, т.к. он происходит также только в растворенном состоянии. Вода служит регулятором t° тела раст., т. е. защищает его от быстрого охлаждения или перегревания. Растения относятся к пойкилотермным организмам, не имеющим постоянной t° тела, которая значительно зависит от t° среды, но в некоторой степени она может регулироваться водой. Этому способствуют такие, свойства воды, как теплоемкость и теплота парообразования. Теплоемкость воды, в сравнении со многими веществами, особенно металлами, очень высока. При нагревании она поглощает много тепла, а при охлаждении выделяет большое его количество. Это приводит к смягчению колебания t° тела растения при изменении t° среды. Также очень высока у воды теплота парообразования. Это приводит к тому, что при ее испарении затрачивается много тепла, которое выделяется органами растения, что вызывает значительное понижение их t°. Физ. Свойства. Плотность воды определяется отношением ее массы к объему при определенной температуре. За единицу плотности воды принята плотность дистиллированной воды при температуре 4°С. Плотность воды зависит от температуры, количества растворенных в ней солей, газов и взвешенных частиц и изменяется от 1 до 1,4 г/см3. Благодаря сильному притяжению между молекулами у воды высокие температуры плавления (0° С) и кипения (100° С). Плотность воды в твердом состоянии меньше, чем в жидком. Следовательно, лед образуется на поверхности водоемов и не опускается на дно. Очень малая теплопроводность. При понижении температуры и давления понижается и теплопроводность. С понижением температуры и понижением плотности уменьшается теплопроводность. Поэтому происходит медленный нагрев и охлаждение водной массы. Проявляется это свойство в том, что снег предохраняет почву от промерзания, а лед - водоемы от промерзания. Формула воды - Н2О (предложена в 1805 г Гумбольдтом и Гей-Люсаком), т.е. состоит из 1 атома кислорода и 2-х атомов водорода. 1). Молекула воды асимметрична, образует равнобедренный треугольник. 2). Молекула воды обладает полярностью, поэтому является электрическим диполем. 3). Молекулярная структура воды: вода находится в трех состояниях и осуществляет фазовые переходы. 2. Поступление воды в растительную клетку. Осмотическое давление и его значение в поглощении воды клеткой. Методы определения осмотического давления Поглощение воды из внешней среды обязательное условие существования любого организма. Вода может поступать в клетку растений благодаря набуханию биоколлоидов, увеличивая степень их гидратации. Такое поступление воды характерно для сухих семян помещенных в воду. Однако главный способ поступления воды в живые клетки является ее осмотическое поглощение. Осмосом называется прохождение растворителя в раствор, отделенный от него полунепроницаемой мембраной(т.е. пропускающей растворитель, но не молекулы растворенных веществ). Природу осмоса стали изучать в 1826 году, когда фран. Физиолог Г.Дютроше сконструировал первый осмометр: пузырь из полупроницаемой пленки(пергамент, животный пузырь) с помещенной в него стеклянной трубочки заполнялся раствором сахара или другого орг. В-ва. После погружения пузыря в чистую водй наблюдался подъем уровня жидкости в трубке. Однако применявшееся пленка не были абсолютно полупроницаемыми и медленно проникавший сахар мешал количественному измерению осмотического давления. В. Пфеффер изготовил «искусственную камеру». Основой ее служил пористый фарфоровый сосуд. Во внутреннюю полость наливался раствор желтой кровяной соли и сосуд помещался в раствор CuSO4. При взаимодействие этих веществ в порах фарфора образовывалось гелеобразная масса железистосинеродистой меди Поступление воды в такого рода осмотическую ячейку приводит к увеличению объема жидкости и поднятию ее уровня в манометрической трубке до тех пор пока гидростатическое давление столба жидкости не повысится настолько, чтобы препятствовать дальнейшему увеличению объема раствора. В достигнутом состоянии равновесия полупроницаемая мембрана в 1 времени пропускает одинаковое количество воды в обоих направлениях. Гидростатическое давление в этом случае соответствует потенциальному осмотическому давлению П* Вант-Гофф: для разбавленных растворов осмотическое давление при постоянной температуре определяется концентрацией частиц (молекул, ионов) растворенного вещества (числом их в 1 объема раствора). Потенциальное осмотическое давление выражается в Паскалях и отражает максимально возможное давление, которое имеет раствор данной концентрации, или максимальную способность раствора в ячейке поглощать воду. П*= i*c*RT, где c-концентрация раствора в молях, T-абсолютная t, R-газовая постоянная, i-изотонический коэффициент, равный 1+α(n-1), где α-степень электролитической диссоциации, n-число ионов, на которые распадается молекула электролита Значение осмотического давления в поглощении воды: от осмот давления зависит сосущая сила клетки, достигая у семян при 6%-ной окружающей влажности величины 4,0510Па(400Атм), что обеспечивает необходимое для прорастания поглощеия воды даже из сравнительно сухой почвы. Осмотическое давление можно определить благодаря фотометрическому методу.
Энергетический уровень молекул данного вещества, который выражается в скорости их диффузии наз-ют химическим потенциалом этого вещества(ψ). Хим потенциал чистой воды называют водным потенциалом. (ψh2o) Он характеризует способность воды диффундировать, испаряться или поглощаться и выражается в Паскалях. Наивысшая величина водного потенциала – у хим. Чистой воды, эта величина принята за нуль. Поэтому водный потенциал любого раствора и биологической жидкости имеет отрицательное значение. Водный потенциал складывается из осмотического потенциала ψs, потенциала давления ψp, гравитационного потенциала ψg, потенциала набухания биоколлоидов. Для оценки степени участия воды в различных химических, биохимических и микробиологических реакциях широко применяют показатель активность воды aw, определяемый как отношение парциального давления паров воды над продуктом к парциальному давлению пара над чистой водой. Показатель «активность воды» был предложен У. Скоттом в 1953 г. и в настоящее время широко применяется на практике. Методы определения водного потенциала: Метод Шардакова основан на подборе раствора, удельный вес, а соответственно и концентрация которого не изменяется после пребывания в нем растительных тканей в течении 20 минут. В этом случае величина осмотического потенциала раствора равна по модулю потенциалу растения.
Так как мембрана избирательно проницаема и вода проходит через нее значительно легче, чем вещества, растворенные в клеточном соке и цитоплазме, при помещении клетки в воду, то по законам осмоса она будет поступать внутрь клетки. Силу с которой вода входит в клетку, называют сосущей силой S. Она тождественна водному потенциалу клетки(ψh2o). Величина сосущей силы определяется осмотическим давлением П* клеточного сока и тургорным (гидростатическим) давлением в клетке(Р), которое равно противодавлению клеточной стенки, возникающему при ее эластическом растяжении. S=П*-Р При замене этих обозначений соответствующими термодинамическими величинами уравнение приобретает следующий вид: -ψh2o=-ψп-ψр В условиях разной оводненности соотношения между всеми компонентами этого уравнения меняются. Когда клетка полностью насыщена водой(полностью тургесцентна), ее сосущая сила равна нулю, а тургорное давление равно потенциальному осмотическому: S=0, П*=Р. Состояние полного тургора наблюдается в клетке при достаточной влажности почвы и воздуха. Если подача воды к клетке уменьшается (при усилении ветра, при недостатке влаги в почве и т.д.), то вначале возникает водный дефицит в клеточной стенке, водный потенциал которых становится ниже, чем в вакуолях, и вода начинает перемещаться в клеточные стенки. Отток воды из вакуолей снижает тургорное давление в клетке и следовательно увеличивает их сосущую силу. При длительном недостатке влаги большинство клеток теряет тургор и растение повядает. В этих условиях Р=0, S=П*. Метод определения сосущей силы: Определение методом полосок (по Лилиенштерн) Принцип метода основан на подборе такой концентрации наружного раствора, при которой погруженные в раствор полоски растительной ткани не меняют своей длины, так как в поступлении воды наступает динамическое равновесие и объем клеток остается неизменным. При более высокой концентрации раствора длина полосок уменьшается. Если осмотическое давление меньше величины сосущей силы клетки, то клетка всасывает воду из раствора, увеличивается в объеме и длина полосок становится больше.
В электрическом поле катиона все ближайшие молекулы воды ориентируются отрицательными полюсами внутрь, а вокруг аниона внутрь направлены положительные полюсы молекулы воды. Этот внутренний, прочно связанный с ионами слой молекул воды называют первичной или ближней гидратацией. В процессе электрофореза он движется вместе с ионом как одно целое. Однако, ион связывая определенное число молекул воды из своего непосредственного окружения, в результате ион-дипольного взаимодействия ориентирует также более далеко расположенные диполи воды. Эту гидратацию называют вторичной(дальней). В растворах содержащих ионы структура воды существенно меняется. В разбавленных растворах (<0,1моль/л) это происходит благодаря заряженным ионам. Маленькие ионы с большей плотностью заряда сильнее действуют на структуру чистой воды по сравнению с большими ионами, имеющими малую плотность заряда, и те и другие разрушают структуру воды: первые притягивают молекулы воды, вторые при внедрении в воду из-за большого размера разрушают льдоподобный каркас. При этом может меняться вязкость водного раствора: структура, создаваемая слабогидратированными большими ионами(с малым зарядом) делает вязкость раствора ниже вязкости чистой воды(Li, Na, Mg,F), а более плотная структура, образуемая гидратированными ионами, обусловливает более высокую вязкость, чем в чистой воде(K, Rb, Cl, OH, NO3) В белках гидратация обусловлена взаимодействиями молекул воды с гидрофильными (ионными и электронейтральными) и гидрофобными (неполярными) группами и ее иммобилизацией в замкнутых пространствах внутри макромолекул при их конформационных перестройках. При ионной гидратации(взаимодействие с -NH3, -СОО группами) и электронейтральной(с -СООН, -ОН, -СО, -NH) молекулы воды электростатически связываются и образуется мономолекулярный слой первичной гидратации. Число ионнизированных групп в белке зависит от рН среды. Наименее гидратирован белок в его изоэлектрической точке, при которой отмечается также самая низкая растворимость белков. Иммобилизованная вода, оказавшаяся замкнутой внутри макромолекул, может участвовать в образовании слоя первичной гидратации, а остальная ее часть сохраняет свойства обычной воды, но с ограниченной подвижностью Свободная вода легко передвигается по растению и испаряется. Она находится в основном в межклетниках, т. е. в свободном пространстве. Связанная вода испаряется и передвигается с трудом. Она находится преимущественно внутри клетки — в цитоплазме и вакуоли. Разделяется на осмотически и коллоидно связанную. Первая соединена с растворенными в ней веществами и находится в вакуоли. Эта связь не очень прочная, и осмотически связанная вода может выходить из клетки, например, при плазмолизе. Свойства коллоидно связанной воды обусловлены наличием белков — коллоидов цитоплазмы, где она и находится. Такая связь большей частью очень прочная, и коллоидно связанная вода выходит из клетки только при очень сильном обезвоживании (продол. засухе).
Вышедшие на сушу растения приспособились к наземному образу жизни благодаря приобретению способности создавать внутри своего тела непрерывный восходящий ток воды. 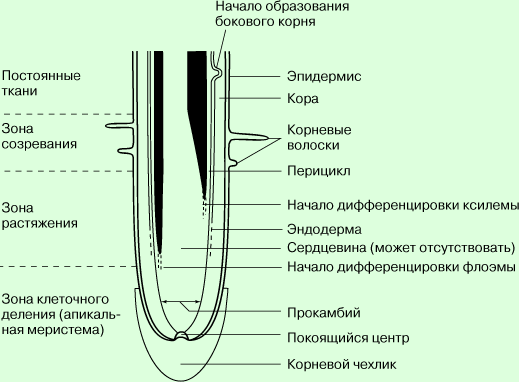 Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растение из почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усиленно делящихся, плотно упакованных клеток, почти целиком заполненных цитоплазмой. Следующая зона — зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а также расстояния от кончика корня, корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков. Некоторое количество воды может поступать и через опробковевшую зону корня. Это главным образом наблюдается у деревьев. В этом случае вода проникает через чечевички. Строение корня в зоне корневых волосков. Поверхность корня в этой зоне покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. Кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системы крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток — эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, пропитывается суберином, что не позволяет передвигаться воде и растворенным в ней веществам. Поступление воды и растворенных веществ в корни через биологические мембраны осуществляется благодаря таким процессам как осмос, диффузия и активный транспорт. Осмос – диффузия воды через мембрану. Диффузия – проникновение веществ через мембрану по градиенту концентрации (из области где их концентрация выше, в область, где их концентрация ниже). Диффузный транспорт веществ воды и ионов осуществляется при участии белков мембраны, в которой имеются молекулярные поры, либо при участии жирорастворимых веществ. Активный транспорт – перенос веществ против их градиента концентрации, связанный с затратами энергии. Он осуществляется специальными белками-переносчиками, которые образуют ионные насосы. Из клеток с корневыми волосками водный раствор просачивается в клеточные поры корня и далее из клетки в клетку попадает в сосуды. По сосудам корня вода с растворенными веществами поднимается в стебель, а по сосудам стебля – почкам, листьям, цветкам. В почве вода находится в разных физических состояниях: жидком, газообразном (водяной пар), твердом (лед), химически и физико-химически связанном с другими веществами и твердыми минеральными, органическими и органо-минеральными частицами. Зависимости от физического состояния и характера связей воды в почвенном среде различают категории, формы и виды грунтовой воды. В почве выделяют следующие категории воды: • химически связанная вода, входящая в состав других веществ (например, гипса) и недоступна для использования растениями; • жесткая вода (лед) - находится в этом состоянии по низкой (отрицательной) температуры и недоступна для растений, однако становится доступной после таяния; • водяной пар, содержащийся в почвенном воздухе и после конденсации становится доступной для растений; • прочно связанная вода, которая содержится адсорбционными силами на поверхности частиц почвы в виде пленки толщиной в два-три диаметра молекул воды, находится в газообразном состоянии (водяной пар) и недоступна для растений; • непрочно связанная вода, которая представляет собой пленки влаги вокруг частичек грунта толщиной до 10 диаметров молекул воды, перемещается между грунтовыми частицами под воздействием сорбционных сил и является труднодоступной для растений; • свободная вода, которая не связана молекулярными силами с частицами почвы, поэтому свободно или под влиянием менисковых сил движется в грунтовых порах и доступна для растений.
Корневое давление-давление возникающее в проводящих сосудах корней растений. К. д. и Транспирация вызываютподнятие пасоки (воды и растворённых в ней питательных веществ) вверх по стеблю. В основе К. д. лежитявление Осмоса; клетки корня активно выделяют в сосуды минеральные и органические вещества, что исоздаёт более высокое, чем в почвенном растворе, Осмотическое давление. К. д. обычно равно 1-3 ат (внекоторых случаях до 10 ат; 1 ат = 105 н/м2) и зависит от условий жизнедеятельности корня. При низкойтемпературе или недостатке кислорода К. д. снижается, при гибели корня равно нулю. Максимум К. д.отмечается днём, минимум — ночью. Плач растений. — Под этим названием известно выделение водянистого сока из древесины растений при поранении их, напр., при просверливании отверстий в стволе или при отрезании ветвей или целого стебля. «Плач» находится в связи с током почвенной воды, поднимающимся по древесине. Особенно энергично П. происходит весною, до распускания почек; (так наз. весенний П.). В это время с пробуждением жизни в растении начинается поднятие воды, а между тем, при отсутствии листьев испарение ее еще очень слабо. Растение наливается поэтому водою, и если в это время просверлить в стволе, напр., березы или клена, отверстие, то из него в изобилии будет выделяться водянистый сок — пасока. Выделение ее происходит со значительною силою, различною, смотря по растению и условиям. По химическому составу пасока представляет водный раствор различных органических (плодовый или тростниковый сахар, белки, органические кислоты) и минеральных веществ. Присутствие этих веществ объясняется тем, что на зиму древесные породы наполняются питательными запасами, которые весною подвергаются растворению и вместе с почвенною водою, всасываемою корнями, передвигаются по древесине в развивающиеся почки. Если же удалить всю облиственную (испаряющую) часть растения, срезав его под корень, то пенек сначала всасывает воду, но через некоторое время, если почва достаточно влажна, начинает выделять ее. Это так назыв. летний П. Летняя пасока не содержит уже органических веществ и представляет чистый раствор почвенных минеральных солей. Гуттация выделение листьями растений капельножидкой воды. Происходит, когда корнями поглощается водыбольше, чем испаряется листьями. Г. наблюдается большей частью ночью, рано утром или при помещениирастений во влажную атмосферу; часто отмечается у молодых проростков, например злаков, развитиекорневой системы у которых обгоняет развитие испаряющей поверхности листьев. Капельки водывыделяются через Водяные устьица под воздействием корневого давления, с силойнагнетающего воду в стебли и листья. Г. -физиологический процесс, связанный с жизнедеятельностьюрастения: если проростки пшеницы поместить под колпак, внутри которого находятся пары хлороформа, тоГ. прекращается. Выделения при Г. всегда содержат ряд минеральных веществ. Очевидно, Г. освобождаетрастение от избытка солей, главным образом кальция. Выделение жидкости идет через специальные водные устьица – гидаторы.
Значения: Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете; Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое; С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом чем интенсивнее транспирация, тем быстрее идет этот процесс.Основным транспирирующим органом является лист. Средняя толщина листа составляет 100—200 мкм. Паренхимные клетки листа расположены рыхло, между ними имеется система межклетников. Эпидермис — покровная ткань листа, состоит из компактно расположенных клеток, наружные стенки которых утолщены. Кроме того, листья большинства растений покрыты кутикулой. Удаление кутикулы во много раз повышает интенсивность испарения. Для соприкосновения листа с атмосферой имеются поры — устьица. Устьице — это отверстие (щель), ограниченная двумя замыкающими клетками. Каждая замыкающая клетка устьица в отличие от клеток эпидермиса имеет хлоропласта. В них происходит фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла. Устьица — одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу клеточные стенки более толстые, а внешние — более тонкие. 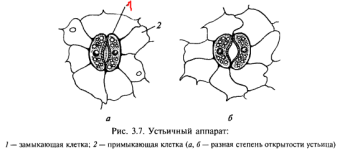 Кутикулярная транспирация Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации.Интенсивность кутикулярной транспирации варьирует у разных видов растений. У молодых листьев с тонкой кутикулой она может составлять около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация равна 1/10 общей транспирации. В стареющих листьях из-за повреждения кутикулы она может возрастать. Таким образом, кутикулярная транспирация регулируется главным образом толщиной и целостностью кутикулы и других защитных покровных слоев на поверхности листьев. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом. Устьичная транспирация Основная часть воды испаряется через устьица. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно Количественные показатели транспирации: Интенсивность транспирации – это количество, г, воды, испаряемой растением в единицу времени (ч) с единицы поверхности (дм 2 ). При определении продукционных характеристик рассчитывают ко- личество воды, израсходованной растением за весь вегетационный период, и относят его к сухой массе всего растения. Транспирационный коэффициент – это количество воды (г), расходуе- мой растением на образование 1 г сухого вещества. Сутчный ход транспирации. У всех растений наблюдается периодичность суточного хода транспирации. У деревьев, теневых растений злаков (гидростабильные виды) испарение воды достигает максимума до наступления максимума дневной температуры. В полуденные часы транспирация падает. Вечером, при снижении дневных температур транспирация снова увеличивается. Такой ход транспирации приводит к незначительным изменениям осмотического давления и содержания воды в клетках в течение дня. У видов, способных переносить резкие изменения содержания воды в клетках в течение дня, транспирация повышается в полдень и падает ночью (гидролабильные виды) Закрывание устьиц в полдень может быть вызвано увеличением уровня углекислого газа в листьях при повышении температуры воздуха (усиление дыхания и фотодыхания), а также возможным водным дефицитом, возникающим в тканях при высоких температурах и низкой влажности воздуха. Это приводит к повышению концентрации АБК и закрыванию устьиц. На поступление воды в растение оказывают влияние внешние условия. 1. Температура. Поступление воды в растение зависит от температуры. С понижением температуры скорость поступления воды сокращается. Это может происходить в результате следующих причин: а) повышается вязкость воды и снижается ее подвижность; б) Тормозится рост корней; в) Уменьшается скорость метаболических процессов; 2. Снижение аэрации почвы (повышение углекислого газа) Повышение концентрации углекислого газа приводит к повышению вязкости воды и снижает проницаемость цитоплазмы. 3. Содержание воды в почве, концентрация почвенного раствора. Вода поступает в корень, если водный потенциал корня ниже, чем водный потенциал почвы. На засоленных почвах или на почвах, где концентрация почвенного раствора очень высокая, водный потенциал почвы ниже. Поэтому вода начнет выделяться из корня. У растений, произрастающих на этих почвах - галофитах, в процессе эволюции выработался такой приспособительный признак как высокая концентрация клеточного сока. Это обуславливает более низкий водный потенциал клеточного сока, вследствие чего вода из почвенного раствора поступает в корни. 9. Устьичная транспирация. Регуляция устьичных движений при действие внешних и внутренних факторов Устьичная транспирация Основная часть воды испаряется через устьица. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно Регуляция устьичной активности. Открывание и закрывание устьиц регулируется несколькими механизмами. 1.Изменение тургора замыкающих клеток. Замыкающие клетки устьиц имеют неоднородные клеточные стенки. Клеточная стенка, которая ближе к устьичнойщели толще, чем противоположная стенка. Целлюлозные волокна располагаются в ней так, что эта стенка менее эластична. По мере того, как клетка поглощает воду, тонкая и эластичная часть клеточной стенки, удаленной от устьичной щели растягивается. Менее эластичный участок клеточной стенки, окаймляющий устьичную щель, растягивается слабее. Поэтому замыкающие клетки устьиц принимают круглую форму и между замыкающими клетками образуется отверстие – устьица раскрываются. Когда вода выходит из замыкающих клеток, то устьица закрываются. 2 Степень обеспеченности клеток водой. Гидропассивная устьичная реакция происходит тогда, когда клетки эпидермиса и хлоренхимы, окружающие устьица, после обильного полива переполняются водой и механически сжимают замыкающие клетки. Если окружающие клетки теряют воду, то устьица открываются. Гидроактивная устьичная реакция. Гидроактивное закрывание устьиц происходит в результате того, что транспирация превышает поглощение воды корнем. При этом тургорное давление замыкающих клетках достигает критического уровня. По мере развития водного дефицита в клетках листа увеличивается содержание фитогормонаабсцизовой кислоты (АБК), которое приводит к снижению тургора клеток, и устьица закрываются. 3.Фотоактивное открывание устьиц. При хорошем освещении, чем выше уровень освещенности, тем шире открываются устьица. Установлено, что днем на свету в замыкающих клетках увеличивается содержание сахаров и, соответственно, повышается осмотическое давление. В замыкающих клетках увеличивается сосущая сила и в них поступает вода. Это приводит к открыванию устьиц. Кроме того, днем на свету взамыкающих клетках накапливаются и ионы калия, что также приводит к повышению осмотического давления и открыванию устьиц. 10. Пути и механизмы передвижения воды по растению Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя(транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала. Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.К такому же выводу можно прийти и при рассмотрении явления плач растенийЕсли срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Передвижение воды по растению Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилем. В 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству. Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа. Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Вся вода в клетке находится в равновесном состоянии. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы. 11. Особенности водного обмена у растений различных экологических групп. Физиологические основы орошаемого земледелия Растения, обитающие в воде - гидратофиты или гидрофиты, погружены в воду полностью или частично. Они регулируют постоянство состава внутренней среды с помощью механизмов защиты от избыточного поступления воды. Первичными гидрофитами являются водоросли. Водные цветковые растения - это вторичные гидрофиты, происходящие от наземных форм. По способности приспосабливать водный обмен к колебаниям водоснабжения различают две группы наземных растений: пойкилогидрические и гомойгидрические. Пойкилогидрические организмы (бактерии, синезеленые водоросли, низшие зеленые водоросли, грибы, лишайники и другие) приспособились переносить значительный недостаток воды без потери жизнеспособности. При этом у них снижается интенсивность обмена веществ, клетки равномерно сжимаются. Протопласт их клеток при сильном обезвоживании переходит в состояние геля. Увеличение количества воды в среде приводит к возобновлению активного метаболизма в клетках. По характеру изменения таких показателей водного режима, как интенсивность транспирации, осмотическое давление, содержание воды в течение суток они относятся к гидролабильным растениям, так как у них значительно изменяются содержание воды и испарение. Гомойгидрические растения (наземные папоротникообразные, голосеменные, цветковые) составляют большинство обитателей суши. Они обладают механизмами регуляции устьичной транспирации, а также корневой системой, обеспечивающей доставку воды. Поэтому даже при значительных изменениях влажности среды у этих растений не наблюдается резких колебаний содержания воды в клетках, в которых, как правило, развита вакуолярная система. Их клетки не способны к обратимому высыханию. У этих растений гидростабильный тип водного режима. Стабилизации водного режима у многих видов растений способствуют запасы воды в корнях, стеблях и запасающих органах. Гомойгидрические растения делятся на три экологические группы: 1. Гигрофиты (тонколистные папоротники, некоторые фиалки и другие), произрастающие в условиях повышенной влажности и недостаточной освещенности. Теневыносливые гигрофиты, с почти всегда открытыми устьицами, имеют гидатоды, через которые выделяют избыток воды в капельножидком состоянии. Гигрофиты плохо переносят почвенную и воздушную засуху. 2. Мезофиты (лиственные деревья, лесные и луговые травы, большинство культурных растений) обитают в среде со средним уровнем обеспеченности водой и не имеют ясно выраженных приспособлений к избытку или недостатку воды. 3. Ксерофиты живут в местах с жарким и сухим климатом и приспособлены к перенесению атмосферной и почвенной засухи. Если годовые осадки превышают годовое испарение, то говорят о гумидной зоне, в обратном случае об аридной. Многие с/х районы расположены в аридной зоне, где земледелие возможно только благодаря искусственному орошению. Орошение — это искусственное увлажнение почвы для получения высоких и устойчивых урожаев сельскохозяй ственных культур. Для подачи воды на поля строят оро сительные системы. Увлажнительное орошение создает в почве нужный водный и воздушный режимы. Оно нужно везде, где ра стения не обеспечены требуемым количеством воды в те чение всего вегетационного периода или его части. Этот вид орошения является основным в России и других стра нах. Удобрительное орошение применяют для внесения удобрения в почву с помощью воды, которая, являясь растворителем удобрений, транспортирует их в увлажня емый слой почвы Разработаны методы определения сроков полива исходя из состояния растения: уровня оводненности листьев, их осмотического давления, степени отверстности устьиц и способности выделять пасоку. Наибольшее распространение получил метод определения сосущей силы листьев, коррелирующийся с тем состоянием водного режима растений, которое еще не вызывает нарушений обмена веществ, и проведенный вовремя полив обычно приводит к повышению урожая. |