ЛЕКЦИЯ 11_Водный обмен. Водный обмен растений
 Скачать 2.64 Mb. Скачать 2.64 Mb.
|

|
ЛЕКЦИЯ 11 ТЕМА: ВОДНЫЙ ОБМЕН РАСТЕНИЙ 1. Значение, структура и свойства воды. 2. Водный обмен клетки. Формы воды в клетке. 3. Механизмы поступления воды в растительную клетку: осмотический, коллоидно-химический, аквапорины. Для процессов жизнедеятельности растений главными экологическими факторами являются: 1) энергия солнечного света; 2) наличие в атмосфере углекислого газа; 3) наличие в атмосфере кислорода; 4) наличие в окружающей среде растворимых минеральных веществ; 5) наличие в окружающей среде воды. Размножение и существование в океане организмов ограничивалось главным образом наличием в окружающей среде необходимых минеральных веществ. В связи с эти автотрофные организмы в процессе эволюции более интенсивно начали развитие у берега. В прибрежной зоне был ряд преимуществ, которые способствовали эволюционному развитию автотрофных организмов: 1) вода обогащена минеральными солями, которые приносятся с суши; 2) действует гораздо больше экологических факторов, под влиянием которых организмы эволюционируют в сторону усложнения структуры и увеличения разнообразия. Ограничивающим фактором распространения растений на суше являлась вода. В связи с этим эволюционная стратегия растений заключалась в следующем: 1) развитие корневой системы – специального органа, который обеспечивал поглощение воды, минеральных веществ и закрепление в грунте; 2) развитие стебля – опоры для листьев и обеспечения непрерывного потока воды и минеральных солей от корня к листьям; 3) покрытие всех надземных органов кутикулой, снижающей потерю воды; 4) развитие специальных отверстий – устьиц, обеспечивающих газообмен между окружающей средой и растением, перемещение воды вверх по растению и поддержание нужного соотношения между расходованием и потреблением воды; 5) формирование в организме специальных тканей, сохраняющих эмбриональное состояние (меристем), которые обеспечивают рост растения в течение всей жизни, что позволяет отыскивать источники воды и минеральных веществ и постоянный приток квантов света; 6) развитие специальных репродуктивных органов, обеспечивающих размножение растений в условиях суши. 1. Значение, структура и свойства воды Вода играет огромную роль в жизни растения. Прямо или косвенно она участвует во всех жизненных процессах, протекающих в клетке. В среднем вода составляет 80–90% массы растения. Вода в биологических объектах выполняет следующие функции: 1. Водная среда объединяет все части организма в единое целое, представляя собой непрерывную среду. Прозрачность водных растворов – это доступность солнечных лучей. 2. Вода – важнейший растворитель и важнейшая среда для биохимических реакций. 3. Вода участвует в упорядочении структур в клетках. Входит в состав белков, определяя их конформацию. Удаление воды из белков приводит их к коагуляции и выпадению в осадок. 4. Вода – метаболит и непосредственный компонент биохимических процессов. В фотосинтезе вода является донором электронов. В дыхании (цикл Кребса) вода принимает участие в окислительных процессах. Она необходима для гидролиза и многих синтетических процессов. 5. Вода обладает относительно высокой протонной и электронной проводимостью, что необходимо для мембранных процессов. 6. Вода – главный компонент в транспортной системе высших растений – в сосудах ксилемы и в ситовидных трубках флоэмы. 7. Вода – терморегулирующий фактор: защищает ткани от резких колебаний температуры благодаря высокой теплоемкости. 8. Вода поддерживает структуру растения в пространстве и является хорошим амортизатором при механических воздействиях на организм, т.к. малосжимаема. Структура воды Вода может находиться в 3-х агрегатных состояниях: газообразном, жидком, твердом (рис. 1). 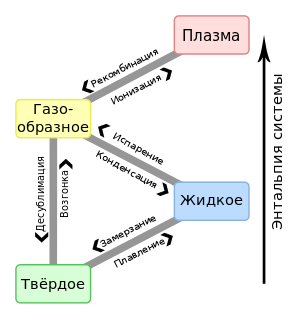 Рис. 1. Агрегатное состояние вещества В каждом из этих состояний структура воды неодинакова. В зависимости от состава находящихся в ней веществ вода приобретает новые свойства. Твердое состояние воды бывает 2-х типов: 1) кристаллическое – лед; 2) некристаллическое – стеклообразное, аморфное (состояние витрификации). При мгновенном замораживании с помощью жидкого азота молекулы не успевают перестроиться в кристаллическую решетку, и вода приобретает твердое стеклообразное состояние. Такое состояние позволяет замораживать живые организмы без повреждений. Физические свойств воды Вода – самое аномальное вещество, хотя принята за эталон меры плотности и объема для других веществ (рис. 2). 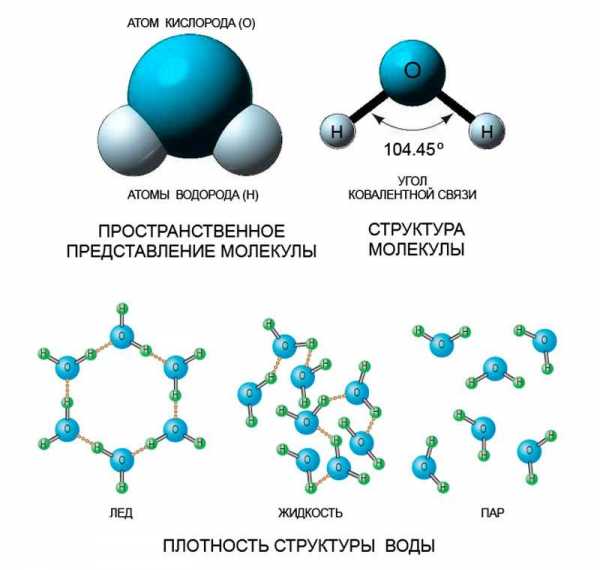  Проекция на плоскости Диполь Рис. 2. Строение молекулы воды 1. Плотность. Все вещества увеличивают свой объем при нагревании, уменьшая при этом плотность. У воды, однако, при давлении в 1 атм в интервале 0–4ºС при увеличении температуры объем уменьшается, и максимальная плотность наблюдается при 4ºС (при этой температуре 1 см3 воды имеет массу 1 г). При замерзании объем воды резко возрастает до 11%. А с таянием льда при 0ºС также резко уменьшается. При 4ºС плотность воды равна 1. 2. Точки замерзания (плавления) и кипения. При давлении 1 атм точки замерзания и кипения воды находятся при 0 и 100ºС, хотя вода относится к гидридам, и температура кипения должна быть около 50–60ºС. Температура кипения воды возрастает с увеличением давления, а температура замерзания (плавления) – падает. 3. Теплота плавления. Скрытая теплота плавления льда очень высока – около 335 Дж/г (для Fe – 25, для S – 40). Служит важным фактором в обороте тепла на Земле. Так, чтобы растопить 1 кг льда или снега, нужно столько же тепла, сколько требуется, чтобы нагреть литр воды до 80ºС. Скрытая теплота парообразования воды – 2,3 кДж/г, это почти в 7 раз выше скрытой теплоты плавления. 4. Вода обладает высокой теплопроводностью. Величина теплоемкости воды в 5–30 раз выше, чем у других веществ. Лишь водород и аммиак обладают большой теплоемкостью. Удельная теплоемкость воды при 16ºС условно принята за 1 и служит эталоном для других веществ. Высокая теплоемкость воды защищает растения от резкого повышения температуры при высокой температуре воздуха, а высокая теплота парообразования участвует в терморегуляции у растений. Все эти показатели свидетельствуют о сильном притяжении молекул воды между собой, вследствие чего жидкая вода обладает большим внутренним сцеплением (когезией). 5. Поверхностное натяжение и прилипание. На поверхности воды из-за некомпенсированности сцепления (когезия) ее молекул создается поверхностное натяжение, величина которого при 18ºС равна 0,72 мН/см (выше только у ртути – 5 мН/см). Благодаря когезии вода как бы покрыта кожицей и способна заполнять тонкие проводящие пучки. Вода обладает также свойством адгезии (прилипания), которое обнаруживается при ее подъеме против гравитационных сил (процесс передвижения воды по растению). В капиллярах сочетаются силы сцепления молекул воды в пограничном с воздухом слое с её адгезией с материалом стенок капилляра. 6. Высокая диэлектрическая проводимость. Вещества, состоящие из молекул с сильно выраженными полярными свойствами, к которым относится и вода, обычно имеют высокие значения диэлектрической проводимости. Это делает электрические силы между растворенными в воде веществами относительно слабыми. Вода является хорошим растворителем для заряженных частиц и плохим – для неполярных молекул. В воде анионы и катионы какой-либо соли оказываются разъединенными. Гидратные оболочки, окружающие ионы, ограничивают их взаимодействие. Положительно заряженные ионы притягиваются к отрицательно заряженным атомам О2 воды, тогда как ионы, несущие отрицательный заряд притягиваются к положительно заряженному Н+. Одновременно нарушается структура самой воды. При этом, чем крупнее ион, тем это нарушение сильнее. 2. Водный обмен клетки и растения Для нормального функционирования клетка должна быть насыщена водой. Известно, что зеленые части растения содержат 80–90% воды. Содержание воды сильно варьирует у разных видов; зависит от типа ткани; возраста растения и его физиологического состояния; изменяется в течение суток и в течение сезона. Даже небольшие изменения в содержании воды вызывают нарушение физиологических функций. То минимальное количество воды, при котором растение способно поддерживать постоянство своей внутренней среды, получило название гомеостатической воды. Для большинства растений это 45–60%. Вода, содержится в: живых клетках, мертвых сосудах, межклетниках. Во взрослой растительной клетке основная масса воды находится в вакуоли, а на долю цитоплазмы приходится лишь 5%. В клеточных стенках ее содержание трудно определить, но считают, что оно колеблется от 25 до 40%. Оводнённость органелл составляет 65%, цитозоля – 95–98%, в ядре содержится 20–30%, в хлоропластах – 14–20%, в митохондриях – 5–7% всей воды протопласта; мембраны содержат 25–30% воды. Содержание воды в различных органах растений колеблется в довольно широких пределах. Оно изменяется в зависимости от условий внешней среды, возраста и вида растений. Так. содержание воды в листьях салата составляет 93–95%, кукурузы – 75–77%. Количество воды неодинаково в разных органах растений: в листьях подсолнечника воды содержится 80–83%, в стеблях – 87–89%, в корнях – 73–75%. Содержание воды, равное 6–11%, характерно главным образом для воздушно-сухих семян, в которых процессы жизнедеятельности заторможены. Вода содержится в живых клетках, в мертвых элементах ксилемы и в межклетниках. В межклетниках вода находится в парообразном состоянии. Основными испаряющими органами растения являются листья. В связи с этим естественно, что наибольшее количество воды заполняет межклетники листьев. Формы воды в клетке Как известно, молекула воды является диполем: один ее полюс заряжен положительно, а другой – отрицательно. Это влияет на состояние воды в клетке, так как в ней, во-первых, содержится много ионов и во-вторых, положительно и отрицательно заряженные группы атомов входят в состав биополимеров. Благодаря этому в клетках есть связанная и свободная вода. Связанная вода имеет измененные физические свойства, главным образом в результате взаимодействия с неводными компонентами. Условно принимают под связанной водой ту, которая не замерзает при понижении температуры до -10º Формы связанной воды в растении: 1) коллоидно-связанная; 2) осмотически-связанная; 3) капиллярно-связанная. К  оллоидно-связанная вода – включает воду, которая находится внутри коллоидной системы и воду, которая находится на поверхности коллоидов и между ними, а также иммобилизованную воду. Иммобилизация представляет собой механический захват воды при конформационных изменениях макромолекул или их комплексов, при этом вода оказывается заключенной в замкнутом пространстве макромолекулы. оллоидно-связанная вода – включает воду, которая находится внутри коллоидной системы и воду, которая находится на поверхности коллоидов и между ними, а также иммобилизованную воду. Иммобилизация представляет собой механический захват воды при конформационных изменениях макромолекул или их комплексов, при этом вода оказывается заключенной в замкнутом пространстве макромолекулы. Значительное количество коллоидно-связанной воды находится на поверхности фибрилл клеточной стенки, а также в биоколлоидах цитоплазмы и матриксе мембранных структур клетки. Осмотически-связанная вода – связана с ионами или низкомолекулярными веществами (рис. 3). Вода гидратирует растворенные вещества – ионы, молекулы. Вода электростатически связывается и образует мономолекулярный слой первичной гидратации. Вакуолярный сок содержит сахара, органические кислоты и их соли, неорганические катионы и анионы. Эти вещества удерживают воду осмотически.   Рис. 3. Осмотически-связанная вода Коллоидно- и осмотически-связанная вода, образующая оболочки вокруг коллоидов или ионов, получила название гидратационной. Количество гидратационной воды зависит от объема и его физиологического состояния и составляет 20–50% от сухой массы. Общее же содержание воды, например, в листьях растений, в 10 раз превышает количество гидратационной. Свободная вода подвижна, она имеет практически все физико-химические свойства чистой воды, хорошо проникает через клеточные мембраны. Свободная вода вступает в различные биохимические реакции, испаряется в процессе транспирации, замерзает при низких температурах. Формы свободной воды 1. Решеточно-упорядоченная форма – образование водородных связей между диполями, в результате чего возникают короткоживущие агрегаты (тетраэдры: одна молекула воды притягивает к себе еще 4 молекулы). Водородные связи непрерывно возникают и рушатся, время полужизни каждой из них 10-9 с. Такие группы называют «мерцающими кластерами» (от англ. сlaster – группа, скопление) (рис. 4, 5).  Рис. 4. Образование решеточно-упорядоченной формы воды 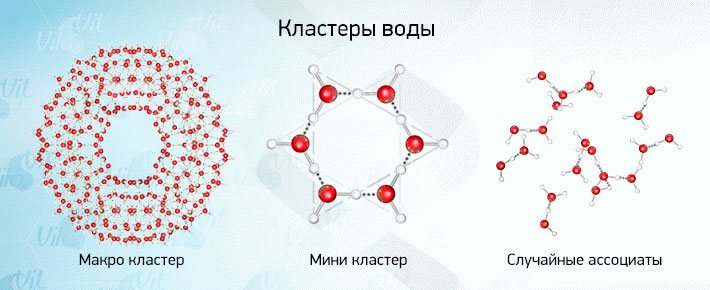 Рис. 5. Кластеры воды 2. Плотноупакованная форма. Тетраэдры соединяются друг с другом и образуется более сложная ажурная решетка – гексагональная структура (рис. 6), в пустотах которой находятся молекулы воды, не образующие никакой структуры. 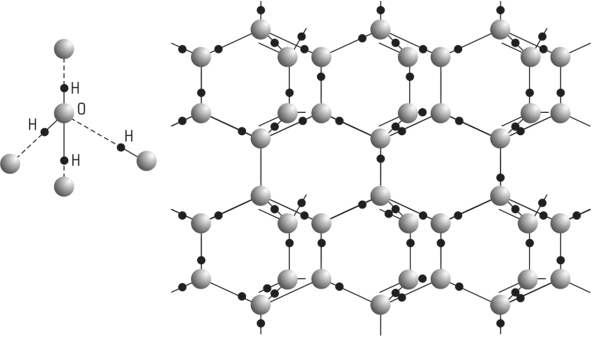 Рис. 6. Гексагональная структура воды Обе фракции воды непрерывно переходят друг в друга. Плотноупакованная вода по своим свойствам напоминает свободную воду. Состояние воды в разных частях клетки неодинаково: 1. В самой цитоплазме находится в основном коллоидно-связанная вода, но есть свободная и осмотически-связанная. 2. В клеточном соке преобладает осмотически-связанная и свободная вода. 3. Клеточные стенки содержат две фракции воды: подвижную и малоподвижную. Капиллярно-связанная вода находится в клеточных стенках и сосудах проводящей системы. Это наиболее подвижная форма, которую относят к категории свободной воды. Значительные количества воды в купных капиллярных пространствах между микрофибриллами целлюлозы легко перемещаются. Примерно треть воды, находящейся в клеточной стенке, связана (гидратирует вещества), остальная часть воды – свободна. Малоподвижная вода – это вода, находящаяся в микрокапиллярных пространствах и та, которая удерживается водородными связями на микрофибриллах целлюлозы. Физиологическое значение свободной и связанной воды различно. Скорость химических реакций, интенсивность физиологических процессов зависят от состояния и структуры вод и, в первую очередь, от содержания свободной воды. 3. Механизмы поступления воды в растительную клетку Как же вода поступает в клетку? В настоящее время говорят о 3-х механизмах: осмотическом, коллоидно-химическом и с помощью аквапоринов. Рассмотрим, как они работают. Осмотический механизм На пути в клетку или органеллу вода, как и другие вещества, должна пройти через плазмалемму, а для поступления в вакуоль – ещё и через тонопласт. Односторонняя диффузия молекул воды или другого растворителя через полупроницаемую мембрану называется осмосом (от греч. оsmos – давление толчок) (рис. 7). 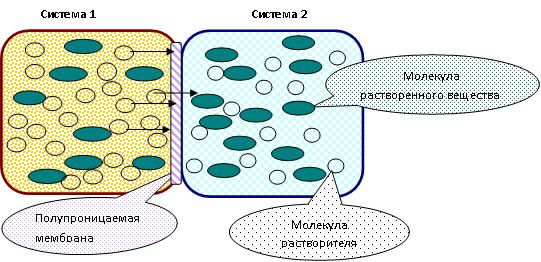 Рис. 7. Осмос Причиной осмоса является разность концентраций растворов по обе стороны полупроницаемой мембраны. В 1748 г. французский аббат А. Ноллет впервые наблюдал, как растворитель проходит через мембрану из разбавленного раствора в более концентрированный. Система, в которой может наблюдаться осмос, называется осмотической. Она состоит из растворов разных концентраций или из раствора и растворителя, разделенных полупроницаемой мембраной. Пространство, окруженное такой мембраной и заполненное каким-нибудь раствором, получило название осмотической ячейки. Изучение осмоса в растительной клетке началось давно. В 1826 г. французский ботаник А. Дютроше сконструировал первый осмометр. В 1877 г. немецкий ботаник В. Пфеффер создал более совершенную модель растительной клетки, названную осмометром Пфеффера. Роль клеточной стенки играл пористый фарфоровый сосуд. Полупроницаемую мембрану получили, налив внутрь фарфорового сосуда раствор медного купороса (CuSO4) и погрузив этот сосуд в другой – с раствором ферроцианида калия (K4[Fe(CN)6]). В результате в порах фарфорового сосуда, где оба раствора соприкасались, возникла полупроницаемая мембрана из ферроцианида меди – Cu2[Fe(CN)6]: 2CuSo4 + K4[Fe(CN)6] → Cu2[Fe(CN)6] + 2K2SO4 Затем фарфоровый сосуд наполнили раствором сахара, играющим роль клеточного сока, и поместили в цилиндр с водой. Вода стала поступать в фарфоровый сосуд. То же самое наблюдается и в клетке, если поместить ее в воду – вода поступает в вакуоль (рис. 8).  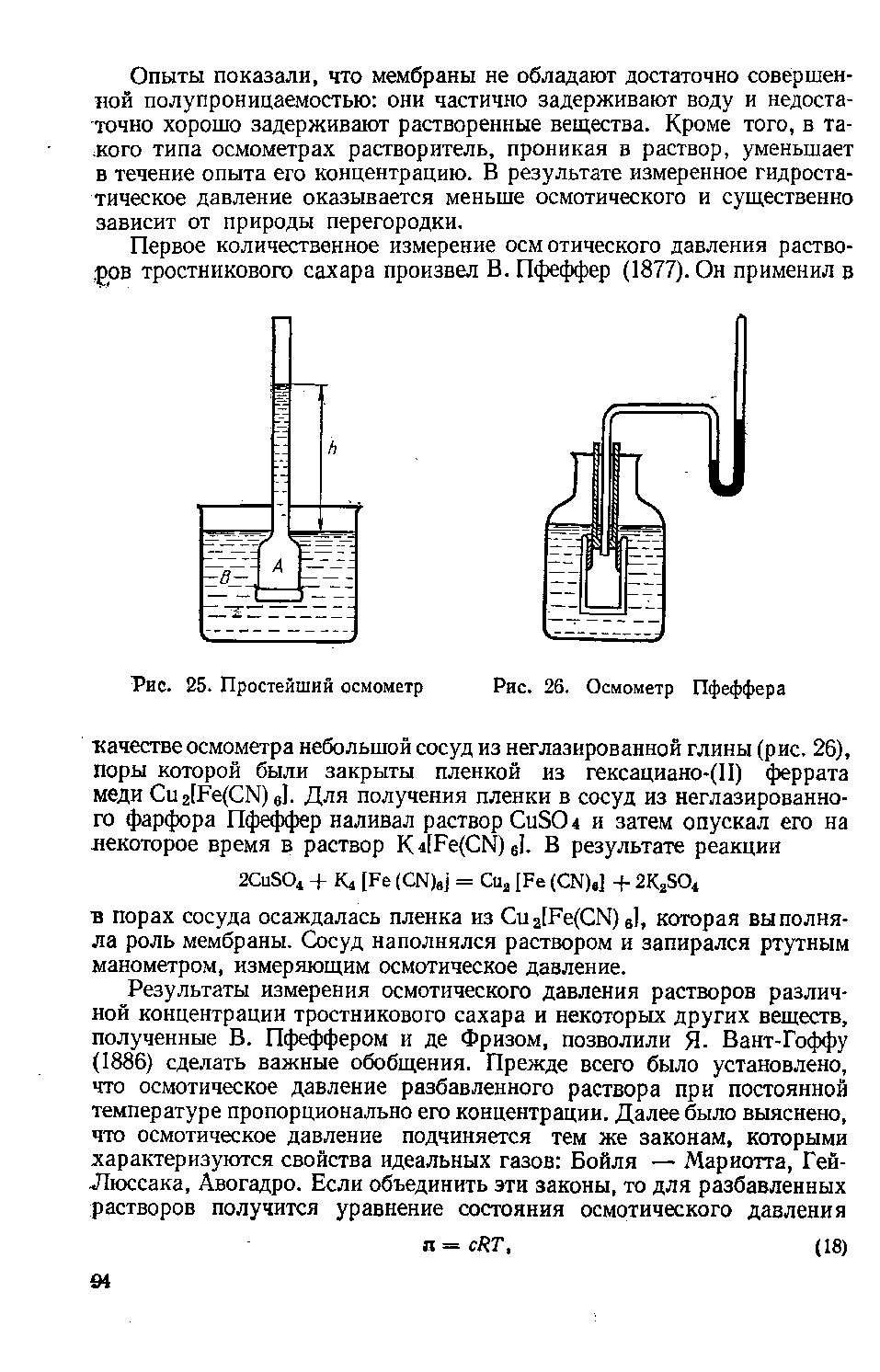 Простейший осмометр Осмометр Пфеффера Рис. 8. Осмометр Пфеффера Таким образом было показано, что клетка представляет собой осмотическую систему (рис. 9). 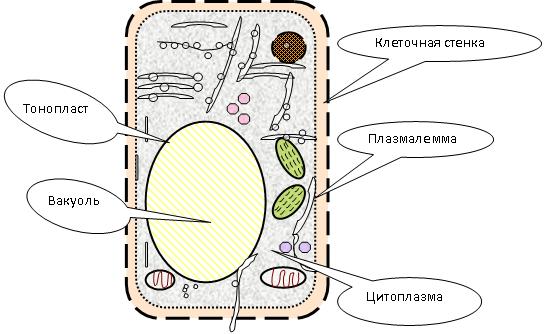 Рис. 9. Клетка как осмотическая система (ячейка) Однако это слишком упрощенное представление о клетке как об осмотической системе. Любая органелла цитоплазмы, окруженная мембраной, представляет собой осмотическую систему. Идеальная полупроницаемая мембрана пропускает молекулы воды и не пропускает молекулы растворенного вещества. Маленькие молекулы воды легко диффундируют через плазмалемму в обоих направлениях: в клетку и из нее. Проницаемость плазмалеммы для воды достаточно велика. Например, если внести тяжелую воду в среду, окружающую корни, то через 1–10 мин процент этой воды в клетках корня будет таким же, как во внешнем растворе. Вещества. Разрыхляющие плазмалемму (например, пипольфен, вытесняющий Са2+ из мембран), увеличивает ее проницаемость для воды и ионов. Как долго вода может поступать в вакуоль? Теоретически поступление воды должно прекратиться, когда концентрация растворов по обе стороны полупроницаемой мембраны сравняется. Однако это не так. В Пфеффер установил, что вода будет подниматься по трубке до тех пор, пока давление водяного столба в ней не станет равным силе, с которой молекулы воды поступают в осмометр. В достигнутом состоянии равновесия полупроницаемая мембрана в единицу времени пропускает в обоих направлениях одинаковые количества воды. То дополнительное давление, которое необходимо приложить к раствору. Чтобы помешать одностороннему току растворителя (воды) в раствор через полупроницаемую мембрану, назвали осмотическим давлением (π). Датский химик Я. Вант-Гофф предложил уравнение для расчета осмотического давления (π): π = RTc, где R – универсальная газовая постоянная (8,314 Дж/(К.моль); T – абсолютная температура (К); с – концентрация раствора в молях. Это уравнение оказалось применимым ко всем разбавленным растворам, кроме растворов электролитов. Электролитическая диссоциация приводит к образованию в растворе большого числа частиц растворенного вещества и этим вызывает увеличение осмотического давления. Поэтому в вышеуказанное уравнение был введен показатель ί (изотонический коэффициент, равный 1 + α (n – 1), где α – степень электролитической диссоциации, n – число ионов, на которое распадается молекула электролита). В результате уравнение расчета осмотического давления приняло вид: π = RTcί Таким образом, осмотическое давление разбавленного раствора при постоянной температуре определяется количеством молекул. Ионов растворенного вещества в единице объема. На величину осмотического давления влияет концентрация только растворенных в воде веществ. Эти вещества называются осмотически активными (осмотиками). К ним относятся органические кислоты, аминокислоты, сахара, соли. Суммарная концентрация этих веществ в клеточном соке варьирует у большинства клеток от 0,2 до 0,8 М. Осмотическое давление выражается в атмосферах, барах или паскалях: 1 атм = 1,013 бара = 105 Па; 103 Па = 1 кПа; 106 Па = 1 МПа Различают 3 вида растворов: 1) изотонические (изоосмотические) – растворы с одинаковым осмотическим давлением; между ними осмос не наблюдается. 2) гипертонические – растворы, имеющие большое осмотическое давление. 3) гипотонические – растворы с малым осмотическим давлением (рис. 10). 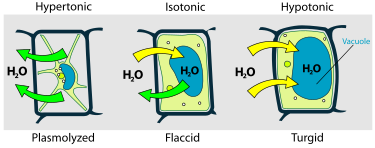 Рис. 10. Движение воды в зависимости от концентрации раствора В 1918 г. немецкие исследователи А Уршпрунг и Г. Блюм доказали, что поступление воды в клетку зависит не только от разности осмотического давления в различных компартментах клетки. Поступая в клетку, вода увеличивает объем вакуоли, которая давит на цитоплазму и заставляет протопласт прижиматься к клеточной стенке. Клеточная стенка растягивается, отчего клетка переходит в напряженное состояние – тургор. Давление протопласта на клеточную стенку называется тургорным. Если бы клеточная стенка могла растягиваться неограниченно, то поступление воды в вакуоль шло бы до тех пор, пока концентрация растворов снаружи и внутри клетки не сравнялась. Но так как клеточная стенка имеет небольшую эластичность, она начинает давить на протопласт в противоположном направлении. Это давление клеточной стенки на протопласт называется тургорным натяжением. Тургорное натяжение в соответствии с третьим законом Ньютона равно по абсолютной величине тургорному давлению, но противоположно ему по знаку. Давление клеточной стенки на протопласт противодействует дальнейшему поступлению воды в клетку. Когда оно станет равным осмотическому давлению, поступление воды в клетку прекратится. Таким образом, осмотическое поступление воды приводит к возникновению гидростатического (тургорного) давления. Разница между осмотическим давлением клеточного сока и противодавлением клеточной стенки определяет поступление воды в клетку в каждый данный момент. Ситуации тургора 1) тургесцентное (ψкл = 0 бар); 2) частичнотургесцентное (ψкл = -10 бар); 3) вялое (ψкл = -20 бар). Явления потери тургора: 1. Плазмолиз – уменьшение объема протопластов и их отделение от клеточных стенок. Внешне клетки и ткани не изменяются (рис. 11, 12). 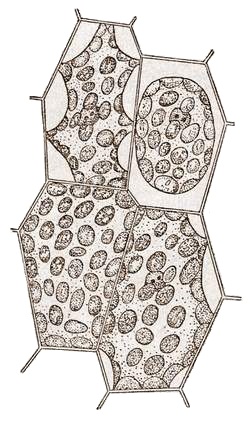 Рис. 11. Плазмолиз растительных клеток 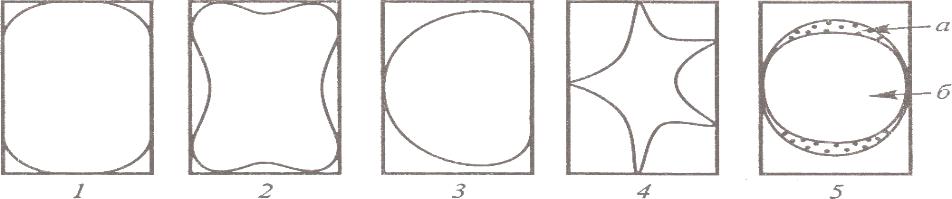 Рис. 12. Формы плазмолиза: 1 – уголковый; 2 – вогнутый; 3 – выпуклый; 4 – судорожный; 5 – колпачковый (а – цитоплазма; б – вакуоль) Отделение протопласта клетки от оболочки происходит под действием на клетку гипертонического раствора. В нормальных условиях плазмалемма растительной клетки плотно прижата к клеточной стенке изнутри под действием тургорного давления. При помещении клетки в раствор, концентрация осмотически активных веществ в котором больше, чем в клеточном соке, – скорость диффузии воды из клеточного сока будет превышать скорость ее диффузии в клетку из вне. Вследствие выхода воды из клетки объем клеточного сока сокращается, тургор уменьшается. Уменьшение объема клеточной вакуоли сопровождается отделением цитоплазмы от оболочки. В процессе плазмолиза протопласт теряет воду, уменьшается в размерах и отделяется от клеточной стенки. 2. Циторриз – протопласты, сокращаясь в объеме, не отделяются от клеточных стенок, а тянут их вслед за собой. В результате клетки и ткани сжимаются. Если поместить взрослые клетки растений (в составе ткани, к примеру, эпидермиса) в гипотонические условия, они не лопнут, поскольку каждая клетка растения окружена более или менее толстой клеточной стенкой. Она служит ригидной структурой, не позволяющей притекающей воде разорвать клетку. Если бы клеточная стенка и плазматическая мембрана клетки могли растягиваться, вода входила бы в клетку до тех пор, пока концентрация осмотически активных веществ снаружи и внутри клетки не выровнялась бы. В реальности клеточная стенка — прочная нерастяжимая структура, и в гипотонических условиях входящая в клетку вода давит на клеточную стенку, плотно прижимая к ней плазмалемму. Клетки растений обычно находятся в гипотонических условиях, поскольку их содержимое богато осмотически активными веществами, большая часть которых (органические кислоты, сахара, соль, низкомолекулярные пигменты) входят в состав клеточного сока, расположенного внутри вакуоли. Тургор – показатель оводненности и состояния водного режима растений. Снижением тургора сопровождаются процессы, увядания и старения клеток. За счет тургора органы растений находятся в выпрямленном, упругом состоянии. Увядание растения – понижение тургорного давления его клеток. Водный потенциал клетки Возможность молекул воды перемещаться из одного места в другое измеряется водным потенциалом (ψВ). Согласно второму закону термодинамики вода всегда движется из области с более высоким водным потенциалом в область с более низким потенциалом. В 1960 г. был введен термин «водный потенциал клетки». Водный потенциал клетки (ψкл.) – это мера энергии, с которой вода поступает в клетку; показатель термодинамического состояния воды в клетке. Молекулы воды обладают кинетической энергией, в жидкости и водяном паре они беспорядочно движутся. Водный потенциал больше в той системе, где выше концентрация молекул воды и больше их общая кинетическая энергия. Максимальным водным потенциалом обладает чистая (дистиллированная) вода. Водный потенциал такой системы условно принят за нуль. При растворении в воде другого вещества, понижается концентрация воды, уменьшается кинетическая энергия молекул воды, снижается водный потенциал. Во всех растворах водный потенциал ниже, чем у чистой воды, т.е. в стандартных условиях он выражается отрицательной величиной. Количественно это понижение выражается величиной, которая называется осмотическим потенциалом (ψосм = ψπ). Осмотический потенциал – это мера снижения водного потенциала за счет присутствия растворенных веществ. Чем больше в растворе молекул растворенного вещества, тем осмотический потенциал ниже. П  ри поступлении воды в клетку ее размеры увеличиваются, внутри клетки повышается гидростатическое давление, которое заставляет плазмалемму прижиматься к клеточной стенке. Клеточная оболочка, в свою очередь, оказывает противодавление, которое характеризуется потенциалом давления или гидростатическим (тургорным) потенциалом (ψтур = ψр). Он обычно положителен и тем больше, чем больше воды в клетке. ри поступлении воды в клетку ее размеры увеличиваются, внутри клетки повышается гидростатическое давление, которое заставляет плазмалемму прижиматься к клеточной стенке. Клеточная оболочка, в свою очередь, оказывает противодавление, которое характеризуется потенциалом давления или гидростатическим (тургорным) потенциалом (ψтур = ψр). Он обычно положителен и тем больше, чем больше воды в клетке.Таким образом, водный потенциал клетки зависит от концентрации осмотически действующих веществ – осмотического потенциала и от потенциала гидростатического (тургорного) давления. Е  сли вода не давит на клеточную стенку (состояние плазмолиза или увядания), противодавление клеточной стенки равно 0, водный потенциал клетки равен осмотическому: сли вода не давит на клеточную стенку (состояние плазмолиза или увядания), противодавление клеточной стенки равно 0, водный потенциал клетки равен осмотическому:ψкл = ψπ По мере поступления воды в клетку появляется противодавление клеточной оболочки, водный потенциал будет равен разности между осмотическим потенциалом и тургорным потенциалом: ψкл = ψπ + ψр Разница между осмотическим потенциалом клеточного сока и противодавлением клеточной оболочки определяет поступление воды в каждый данный момент. Если клеточная оболочка растягивается до предела, осмотический потенциал целиком уравновешивается противодавлением клеточной оболочки, водный потенциал становится равным нулю, вода в клетку перестает поступать: ψπ = ψр; ψкл = 0 После этого клетка уже не сможет поглощать воду ни из какого раствора, не сможет она и отнять ее от другой клетки. Это состояние наблюдается в клетках при достаточной влажности почвы и воздуха. Т 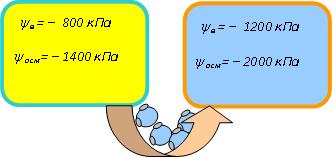 аким образом, вода всегда поступает в сторону более отрицательного водного потенциала: от той системы. Где энергия больше, к той системе, где энергия меньше. аким образом, вода всегда поступает в сторону более отрицательного водного потенциала: от той системы. Где энергия больше, к той системе, где энергия меньше.Значение величины осмотического потенциала имеет большое значение, в частности, для экологических исследований. Его величина позволяет судить о максимальной способности растения получать воду из почвы и удерживать ее, несмотря на засушливые условия. Эта величина колеблется в широких пределах: от -0,1 до - 20 МПа. У большинства растений умеренной зоны осмотический потенциал колеблется от -0,5 до -3,0 Мпа. У пресноводных растений его величина составляет около -0,1 Мпа, у морских водорослей – от -3,6 до -5,5 Мпа. Обычно отрицательная величина осмотического потенциала больше у мелких клеток по сравнению с крупными. Однако даже соседние клетки одной ткани могут различаться по его величине. Величина осмотического потенциала изменяется и в пределах растения: у корней -0,5…- 1,0, у верхних листьев – до -4,0 Мпа. Это обусловливает существование градиента водного потенциала клеток от корней к листьям. У молодых растений осмотический потенциал меньше, чем у старых. У деревьев он отрицательнее, чем у кустарников; у кустарников более отрицателен, чем у трав. Величина осмотического потенциала зависит также от температуры, интенсивности света. Они определяют ее годичные и суточные колебания. Около полудня потеря воды в результате транспирации и накопление продуктов фотосинтеза в клетках листьев вызывают уменьшение осмотического потенциала. У водных растений колебания осмотического потенциала зависят только от скорости фотосинтеза. Растение может регулировать величину осмотического и, следовательно, водного потенциала. Превращение сложных нерастворимых веществ в растворимые (крахмал → сахар, белки → аминокислоты) приводит к возрастанию концентрации клеточного сока и понижению водного потенциала. Однако, осмос – это не просто односторонняя диффузия, для него необходима энергия. Факторы, стимулирующие дыхание, ускоряют поступление воды в клетку и, наоборот, факторы, ингибирующие его, уменьшают ее поступление. Следовательно, для поступления воды в клетки нужна энергия АТФ. Почему для осмоса нужна энергия? Во-первых, нужно иметь растворы разной концентрации по обе стороны мембраны; энергия тратится на активный транспорт растворенных веществ в вакуоль и создание градиента концентраций. Во-вторых, осмотически активные вещества, накапливающиеся в вакуоли, являются продуктами метаболизма, следовательно, для их образования тоже затрачивается энергия. В-третьих, энергия необходима для сохранения избирательной проницаемости мембран. Коллоидно-химический механизм (набухание) Осмотическое поступление воды связано в основном с растворенными веществами клеточного сока, т.е. с вакуолью. Однако около 80% сухой массы протопласта составляют высокомолекулярные соединения: белки, полисахариды, нуклеиновые кислоты, нерастворимые в воде. Они тоже влияют на поступление воды в клетку. Имея положительно и отрицательно заряженные группы атомов (–СОО-, –NH3+), гидрофильные коллоиды притягивают диполи воды. Молекулы воды могут удерживаться около молекулы полимера также с помощью водородных связей, которые возникают между атомами водорода и азота или кислорода. Поглощение жидкости или пара высокомолекулярным веществом, сопровождаемое увеличением его объема, называется набуханием. Набухание рассматривают как особый вид диффузии, т.к. движение воды идет по градиенту концентрации. Силу набухания называют матричным потенциалом (ψматр.). Он зависит от наличия высокомолекулярных компонентов клетки. Может достигать -100Мпа (при поглощении воды сухими семенами). Матричный потенциал всегда отрицательный, т.к. коллоиды, связывая воду, уменьшают ее активность. Поэтому водный потенциал клетки будет равен: ψкл = ψπ + ψр + ψматр. Набухание участвует в поступлении воды в цитоплазму и клеточную стенку. Клеточная стенка способна к набуханию, т.к. имеет в своем составе гемицеллюлозы и пектиновые вещества. Кроме того, набухание клеточной стенки может быть вызвано накоплением воды в межфибриллярных и межмицеллярных пространствах. В вакуоли, как правило, нет набухающих веществ. Некоторые части растений (например, сухие семена) поглощают воду исключительно с помощью набухания. Особенно сильной способностью к набуханию отличаются семена бобовых с высоким содержанием белков. Итак, в зависимости от возраста и строения клетки разные механизмы поступления воды играют бòльшую или меньшую роль. Во взрослой клетке с большой центральной вакуолью главным механизмом является осмотический. в эмбриональной клетке, не имеющей центральной вакуоли, но быстро синтезирующей белковые молекулы, способные притягивать воду, основным механизмом ее поступления становится набухание. В онтогенезе органа может происходить смена механизмов поступления воды. Например, прорастающее семя поглощает воду за счет набухания, а потом включается в работу и осмотический механизм. Аквапорины В настоящее время клетку не рассматривают только как осмотическую систему. Это доказывает открытие аквапоринов. Аквапорины – это специальные белки, которые встраиваясь в мембраны, образуют в липидном бислое водные каналы, или поры. Отсюда и название этих белков (от лат. aqua – вода и poros – отверстие). Молекулярная масса аквапоринов варьирует от 23 до 31 кДа. Молекула такого белка состоит из 6-ти субъединиц, пересекающих липидный бислой. Ее внутренняя поверхность гидрофильна, что позволяет молекулам воды легко «проскакивать» через мембрану (рис. 13).  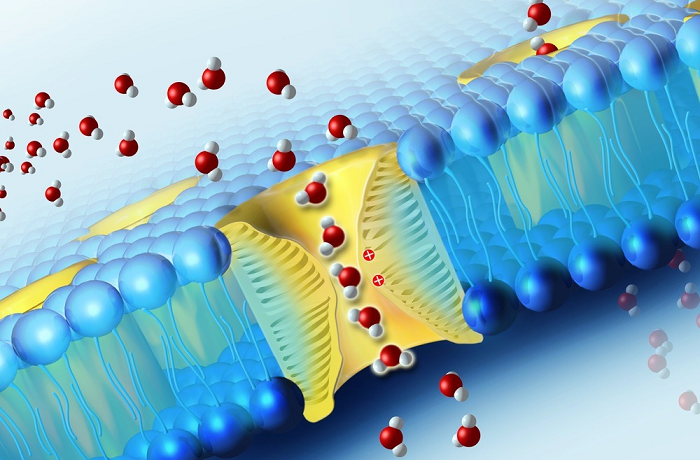 Рис. 13. Аквапорины Аквапорины обеспечивают ток воды через мембрану в обоих направлениях в зависимости от направления градиента водного потенциала, а также отвечают за селективность к молекулам воды. Эти белки найдены практически в клетках всех растений. Удивительной особенностью аквапоринов является их огромное разнообразие. Все аквапорины в зависимости от их локализации в клетке делятся на 2 большие группы: 1) аквапорины плазмалеммы (PIP-белки); 2) аквапорины тонопласта (TIP-белки). Растительные аквапорины обнаружены еще в конце 80-х годов прошлого столетия, но лишь спустя много лет было установлено, что они могут функционировать как очень эффективные водные каналы, ускоряющие диффузию молекул воды. Помимо трансмембранного переноса воды они транспортируют через мембраны маленькие молекулы органических веществ (осмолитов), обеспечивающих осморегуляцию в цитоплазме. Аквапорины не только отвечают за быстрый транспорт воды через мембраны, но также дают организму возможность регулировать водные потоки как между клетками, так и внутри клеток. Эта регуляция осуществляется как путем изменения числа водных каналов в мембране, так и путем изменения их активности. Содержание водных каналов в мембране зависит от биосинтеза аквапоринов, который находится под контролем транскрипционных факторов. Регуляция активности уже сформированных водных каналов осуществляется путем фосфорилирования. Водная проводимость мембран меняется в онтогенезе, изменяя потоки воды в соответствии с запросами роста и развития растения. Установлено, что при росте, когда происходит растяжение клеток, активность генов TIP-белков возрастает. Контролирование водных потоков в растении имеет большое значение и в условиях водного дефицита. ЛЕКЦИЯ № 12 |