ЛЕКЦИЯ 11_Водный обмен. Водный обмен растений
 Скачать 2.64 Mb. Скачать 2.64 Mb.
|

|
ТЕМА: ПОСТУПЛЕНИЕ ВОДЫ В РАСТЕНИЕ 1. Поступление воды в растение. 2. Механизмы поступления воды. 3. Факторы, влияющие на поступление воды. Водные растения, как правило, не испытывают недостатка в воде. Вышедшие на сушу растения приспособились к наземному образу жизни благодаря приобретению способности создавать внутри своего тела непрерывный восходящий ток воды. Этот ток начинается на поглощающей воду поверхности корней, пронизывает все растение и заканчивается на испаряющих поверхностях наземных органов, главным образом, листьев. Причем испарение воды листьями должно быть компенсировано поглощением воды корнями. Таким образом, водный обмен растения состоит из 3 этапов: 1) поступление воды; 2) транспорт воды; 3) выделение воды. Для нормального роста и развития растений необходимо, чтобы расход воды примерно соответствовал приходу, или, иначе говоря, чтобы растение сводило свой водный баланс без большого дефицита. Для этого в растении в процессе естественного отбора выработались следующие приспособления: 1) к поглощению воды – колоссально развитая корневая система; 2) к передвижению воды – специальная проводящая система; 3) к сокращению испарения – система покровных тканей и система автоматически закрывающихся устьиц. 1. Поступление воды в растение Растение поглощает воду 2 способами: 1. Через листья и стебли. Смоченные дождем или росой листья могут поглощать до 25% падающей на них воды. Вода может поступать в листья и непосредственно из окружающего воздуха, влажность которого 100%. Но это поглощение незначительное. 2. Через корни. Основным органом поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности поступление воды происходит из большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой. Общая поверхность корней обычно превышает поверхность надземных органов в 140–150 раз. Так, число корней у 1-летних сеянцев яблони достигает 45 тысяч, у 12–14-месячного растения сахарного тростника общая длина корней может достигать 17 км, а их общая поверхность без корневых волосков – около 400 тысяч см2. Корневые системы однолетних злаков проникают на глубину 1,5–2 м. однако, если считать, что глубина пахотного слоя составляет 20–25 см, то значительная часть корневой системы расположена ниже. Строение корня Из курса ботаники вам известно строение корня. Рассмотрим особенности его анатомии, которые непосредственно влияют на поступление воды. В первичном строении корня различают несколько тканей. Эпиблема (ризодерма) – состоит из одного слоя клеток, покрывает молодые корни. Ее главная функция – поглощение веществ. Наружные стенки клеток эпиблемы не имеют ни кутикулы, ни воска, поэтому они хорошо приспособлены для поглощения воды. Некоторые клетки эпиблемы вытягиваются и превращаются в корневые волоски (рис. 1). Благодаря корневым волоскам поверхность корня, соприкасающаяся с почвой, увеличивается в 10–15 раз. Кроме того, транспортные белки, находящиеся в плазмалемме корневых волосков, активнее, чем в других клетках. Из корневых волосков идет больше плазмодесм в клетки экзодермы. Корневые волоски живут несколько дней и разрушаются. Вместо них в верхней части зоны растяжения образуются новые. Корневой волосок имеет большую центральную вакуоль, поэтому главный механизм поступления в него воды – осмотический. Несмотря на важную роль корневых волосков, у некоторых растений (например, авокадо) они отсутствуют. Их нет и у водных растений. Постепенно, по мере роста корня эпиблема заменяется пробкой. Через опробковевшие части вода практически не поступает в корень. Бòльшую часть (до 80–90%) площади поперечного сечения молодого корня составляет первичная кора. В ее крупных клетках центральные вакуоли занимают около 90% объема протопласта. В состав клеточных стенок входит только целлюлоза, поэтому они хорошо проницаемы для воды. В коре много межклетников, которые в зависимости от условий могут быть заполнены воздухом или водой. Чем толще слой коры, тем больше диаметр корня и тем больше площадь его поверхности, покрытая эпиблемой. Внутри корня находится центральный цилиндр, отделенный от коры эндодермой. Эндодерма состоит из одного слоя клеток. В радиальных и поперечных стенках этих клеток откладывается суберин, а у некоторых растений – лигнин. Образуются так называемые пояски Каспари, непроницаемые для воды и веществ. Это влияет на транспорт воды внутри корня. 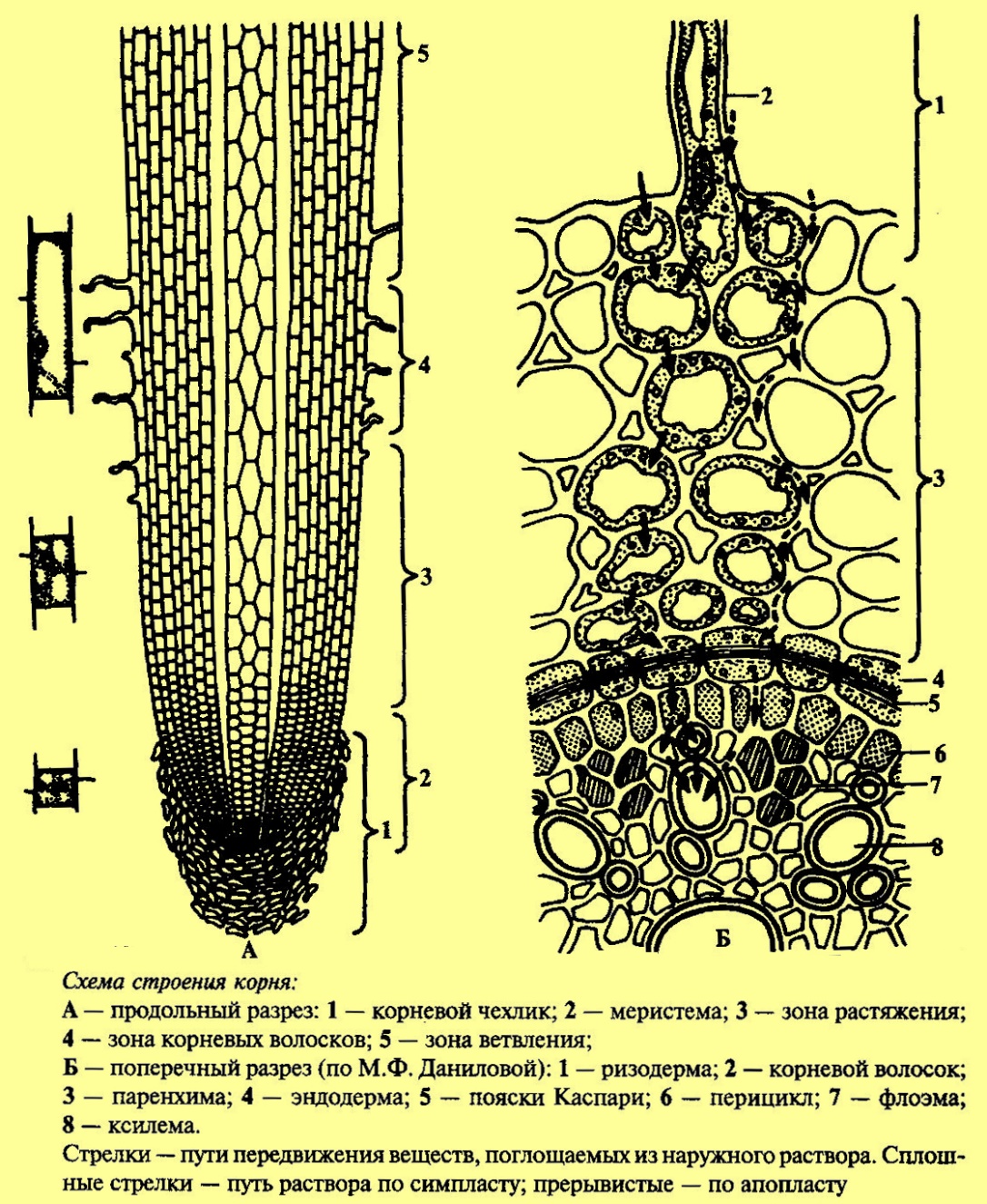 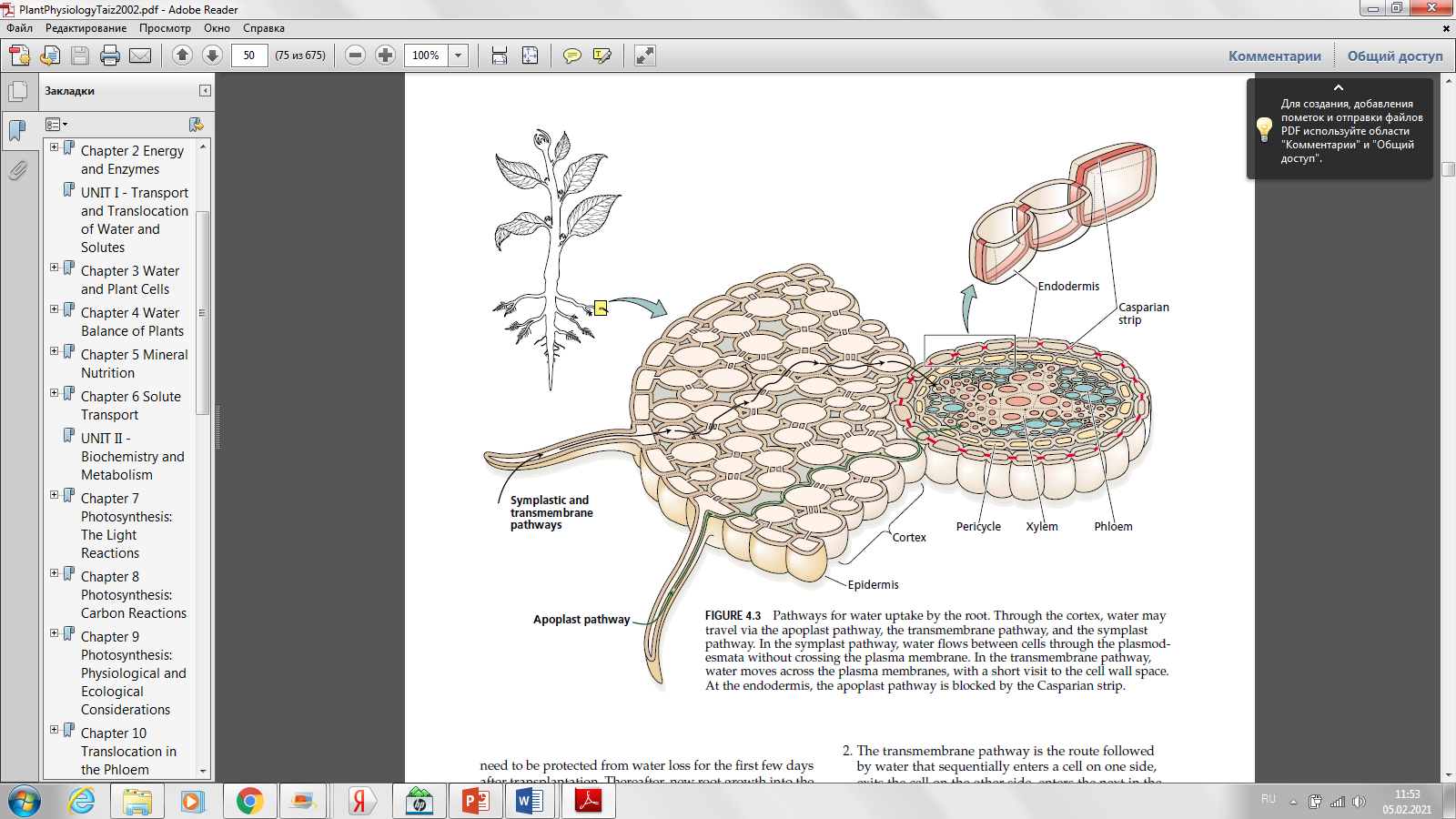 Рис. 1. Строение корня Непосредственно за эндодермой находится перицикл. В состав центрального цилиндра входят проводящие ткани: ксилема и флоэма. Большие сосуды ксилемы окружены живыми клетками ксилемной паренхимы. Крупные поры в клеточных стенках соединяют их с сосудами. Флоэма состоит из ситовидных трубок, трубок-спутниц и флоэмной паренхимы. С помощью проводящих тканей поддерживается связь между корнем и другими частями растения. Все перечисленные ткани образуются из меристематических клеток, находящихся на верхушке, поэтому по длине корня различают 4 зоны: 1) деление клеток (1,5–2 мм от кончика); 2) растяжение клеток (2–7 мм); 3) дифференцировки (корневых волосков); 4) опробковения. Клетки зоны деления защищены корневым чехликом. Основными функциями корневого чехлика являются: 1) предохранение растущей зоны от повреждения при соприкосновении с почвой; 2) облегчение продвижения кончика корня в почве; 3) восприятие геотропического стимула, обеспечивающего правильную ориентацию корней в пространстве. Апикальные клетки меристемы корня (1–2% от общего числа клеток в меристеме) отличаются от остальных более редкими делениями, более медленным синтезом ДНК, РНК и белков, меньшим числом плазмодесм с соседними клетками. Они получили название покоящегося центра. При повреждении апикальной меристемы корня из-за неблагоприятных условий (засуха, механические повреждения и т.д.) фонд ее клеток восстанавливается за счет деления клеток покоящегося центра. За время жизни в меристеме корня каждая клетка делится 6–7 раз. Количество клеток увеличивается вдоль оси корня, в результате чего образуются продольные ряды клеток. Уже в меристеме ткани корня дифференцируются: – формируется перицикл; прослеживается эндодерма, клеточные стенки которой еще лишены субериновых поясков Каспари; – начинается образование элементов флоэмы. Прекратившие деление клетки переходят к растяжению также в направлении оси корня. В зоне растяжения заканчивается дифференцировка флоэмы и формируются элементы протоксилемы – клетки, содержащие цитоплазму с органоидами. В клетках зоны растяжения идет интенсивный синтез белков, о роли которых в процессе набухания мы уже говорили. Дифференцировка тканей корня заканчивается в зоне корневых колосков, где завершается образование основных тканей корня: эпиблемы, первичной коры, эндодермы и системы тканей центрального цилиндра. Вода поступает главным образом в клетки зон растяжения и особенно корневых волосков. Обычно у одного корня длина зоны активного поглощения веществ равняется 5 см. Чем медленнее растет корень, тем больше зон активного поступления воды имеет корневая система растения. Некоторое количество воды может поступать и через опробковевшую часть корня, что характерно, в основном, для деревьев. В этой зоне поступлению воды способствуют и поранения. Вода движется в почве за счет диффузии очень медленно: в течение месяца не более чем на 30 см. Поэтому, в почве не вода движется к корню, а корень движется к воде в процессе роста. В целом корневые системы значительно менее разнообразны по сравнению с надземными органами, в связи с тем, что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем. Большое значение для формирования корневых систем имеет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватывают атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые – за счет усвоения выпадающих осадков. Развитие корневых систем зависит от аэрации. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему. Значение имеют и условия минерального питания. Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению. Особенности корня: 1. Рост является первой и важнейшей особенностью корня как органа. Рост корня, его ветвление продолжается в течение всей жизни растительного организма, т.е. практически он не ограничен. Рост корней отличается большой скоростью. Считается, что одно растение риса в благоприятных условиях может образовать до 5 км новых корней в сутки. За счет этого прироста корневой системы в растение может дополнительно поступать до 1,5 л воды. Только благодаря такому интенсивному росту корневые системы растения могут использовать скудно рассеянную в почве воду. В засушливых условиях формируется в 3–4 раза бòльшая корневая система, чем во влажных. Во время засухи сначала тормозится рост побегов, а потом уже корней. 2. Положительный гидротропизм – при недостатке воды растущие части корня изгибаются и растут в сторону влажных участков почвы. 3. Положительный геотропизм – способность ориентироваться в пространстве. При прорастании семян корень первоначально ориентируется в гравитационном поле, направляя свой рост к центру Земли. Действие гравитационного поля воспринимается статолитами – клетки центральной части корневого чехлика, содержащие много крахмала. Это оказывает влияние на транспорт абсцизовой кислоты (АБК) из кончика корня в зону растяжения. Одностороннее увеличение содержания АБК в зоне растяжения тормозит рост и приводит к целесообразным ростовым изгибам. Избыток индолилуксусной кислоты (ИУК), поступающей из надземной части растяжения, также тормозит удлинение клеток в зоне растяжения, но способствует образованию боковых и придаточных корней. Растущий дистальный конец корня очень чувствителен к механическим давлениям и проникает лишь в рыхлые участки почвы. При сильном механическом сопротивлении рост его в длину тормозится и одновременно в зоне растяжения появляется утолщение (из-за образования стрессового этилена). Это способствует расширению пор в почве и дальнейшему росту корня. Продвижение кончика корня между частицами почвы облегчает корневой чехлик, клетки которого постоянно секретируют во внешнюю среду слизистые вещества (полисахариды). Кроме того, клетки ткани корневого чехлика сами легко отделяются и разрушаются. Это способствует не только продвижению, но и защищает корень от повреждений. При недостатке воды именно она становится главным фактором, определяющим поведение корня. Вместо ориентации в гравитационном поле корень начинает реагировать на градиент влажности. Впервые это было показано Ю. Саксом в прошлом веке в простых опытах. Корни проростков ориентируются первоначально в гравитационном поле и выходят снизу из косо подвешенного влажного блока (сделанного, например, из мха сфагнума). Через некоторое время они изгибаются под острым углом и начинают расти, приближаясь к блоку (где больше влажность). При недостатке воды резко увеличивается поверхность поглощающей зоны за счет интенсивного роста корневых волосков. Таким образом, корни растений удлиняются, чтобы достичь новых участков почвы и развивают большую поверхность соприкосновения с частицами почвенного поглощающего комплекса благодаря ветвлению, образованию все новых и новых поглощающих зон корней и корневых волосков. 2. Механизмы поступления воды Поглощение воды и ее поднятие вверх осуществляется в результате совместного действия следующих факторов: 1) присасывающей силы транспирации, или верхнего концевого двигателя; 2) корневого давления, или нижнего концевого двигателя. 1. Работа верхнего концевого двигателя мало связана с жизнедеятельностью корневой системы. Опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Механизм работы верхнего концевого двигателя несложен и основан на следующем. Вода всегда передвигается в сторону меньшего водного потенциала. Водный потенциал максимален в почве, несколько меньше в клетках корня и самый низкий – в клетках, примыкающих к эпидерме листа. Атмосфера обычно не насыщена водными парами, поэтому обладает самым низким водным потенциалом. При относительной влажности воздуха 90% он равняется -14 МПа. У большинства растений водный потенциал листьев колеблется от -0,1 до -3,0 МПа. Вследствие большой разницы водных потенциалов происходит транспирация. Поскольку концентрация водяных паров в воздухе, даже в условиях влажного климата, как правило, меньше, чем в межклетниках листа, пар движется из тканей растения наружу. Причем диффузия молекул воды от испаряющей поверхности в воздух происходит в 1500 раз быстрее, чем их поступление в корень из почвы. Уменьшение количества воды в паренхимной клетке листа вызывает снижение активности воды в ней и уменьшение водного потенциала. Вода из соседней клетки поступает в паренхимную клетку, транспирирующую активнее. Водный дефицит постепенно от клетки к клетке доходит до корней, и активность воды в них снижается. Вследствие этого вода поступает из почвы в корень. Таким образом, передвижение воды по растению, как и поступление ее в корни, связано с градиентом водного потенциала в системе «почва–растение–атмосфера». Этот градиент тем больше, чем больше клетка листа будет терять воды, т.е. чем сильнее транспирация. Вызванное транспирацией движение воды по растению называют транспирационным током. Транспирационный ток, в свою очередь, обусловливает поступление воды из почвы в растение также по градиенту водного потенциала. Из-за транспирации водный потенциал в верхней части растения ниже, чем у основания. Кроме того, скорость транспирации больше скорости поступления воды, потому что эндодерма и клеточные стенки оказывают сопротивление движению воды. 2. Примером работы нижнего концевого двигателя служит так называемый плач растений и гуттация. Плач растений. Весной у деревьев с еще нераспустившимися листьями можно наблюдать интенсивный ксилемный ток жидкости снизу вверх через надрезы ствола и даже верхних веток кроны (раненое дерево «плачет»). Явление «плача» свидетельствует о значительном корневом давлении, которое в этот период у основания ствола достигает не менее 10 атм (1,013 МПа). Плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток корня. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь, интенсивности дыхания, прекращает «плач». В отсутствие кислорода, под влиянием дыхательных ядов, при пониженной температуре «плач» приостанавливается. У вегетирующих растений при удалении стебля с листьями из оставшегося пенька довольно долго выделяется ксилемный сок, или пасока (вода с растворенными веществами). Корневое давление можно измерить, поместив на пенек манометр. Его величина равна примерно 1–1,5 бара в оптимальных условиях. 2. Гуттацию можно наблюдать на неповрежденном растении. При высокой влажности воздуха на концах и зубчиках листьев выделяется капельножидкая влага – растение гуттирует. Выделение жидкости идет через специальные водяные устьица – гидатоды. Выделяющаяся жидкость – гутта. В ней обнаружены минеральные и органические соединения. Гуттационное выделение влаги листьями особенно характерно для тропических растений, у которых в условиях повышенной влажности транспирация затруднена. «Плач» и гуттация связаны наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую в растении односторонний ток воды с растворенными веществами называют корневым давлением. Итак, корень подает воду в побеги, прежде всего, в стебель, а потеря молекул воды в верхней части водного столба в результате испарения заставляет ее подниматься по сосудам ксилемы для ликвидации этой потери. Поэтому корневое давление было названо нижним, а транспирация – верхним концевым двигателем водного тока. Работают оба двигателя по-разному. Верхний концевой двигатель развивает силу в -1,0…-1,5 МПа и даже более, нижний – в -0,15…-0,3 МПа. Значит, главную роль в транспорте воды играет верхний двигатель. При отсутствии листьев у деревьев весной или после сухого сезона работает только нижний концевой двигатель. Большую роль в поднятии воды по растению нижний концевой двигатель играет также в условиях высокой влажности воздуха, когда транспирация минимальна. Для работы верхнего концевого двигателя источником энергии является солнце, т.к. большая часть поглощаемой световой энергии превращается в тепло и расходуется на транспирацию. Таким образом, верхний концевой двигатель представляет собой автоматически работающий механизм, который тем сильнее «присасывает» воду, чем быстрее ее расходует. Для нижнего концевого двигателя источник энергии – дыхание клеток корней, поставляющих АТФ. Энергия АТФ расходуется на поглощение солей, работу сократительных белков и, следовательно, на создание градиента водного потенциала. Работа обоих концевых двигателей водного тока хорошо согласована. Это доказано рядом опытов П.С. Беликова (СССР, 1968). Если корни растения переносят в питательный раствор, имеющий более низкую температуру (например, 15º вместо 25ºС), то в первые 10 минут наблюдается снижение выделения пасоки почти до нуля, зато интенсивность транспирации возрастает на 30%. Следовательно, ухудшение работы нижнего концевого двигателя усиливает в первые минуты работу верхнего концевого двигателя. С другой стороны, выдерживание в течение часа корней в растворе абсцизовой кислоты (1 мг/л), подавляющей транспирацию, вызывает временное (в первые 10 минут) увеличение выделения пасоки в 2–4 раза. Следовательно, чем слабее транспирация, тем интенсивнее работает нижний концевой двигатель. Такое биологически важное приспособление выглядит, на первый взгляд, парадоксально: поступление воды ухудшилось, лист на этот неблагоприятный фактор отвечает не снижением, а увеличением транспирации. Повышение транспирации в данном случае должно стимулировать поступление воды в корень. Можно предположить, что в растении существует особая регуляторная система – гидродинамическая. Эта система позволяет растению очень быстро реагировать на изменения внешних условий потенциально способных нарушить водный гомеостаз, контролирует водный режим растения, а также ход других функций, например, фотосинтеза. Скорость передвижения воды определяется разностью водных потенциалов в начале и в конце ее пути, а также сопротивлением, которое оно встречает. Она зависит от скорости транспирации и диаметра сосудов. В широких сосудах вода встречает меньшее сопротивление, хотя в них быстрее могут попасть пузырьки воздуха, мешающие движению воды. Скорость движения воды изменяется в течение суток (днем она больше) и отличается у разных растений: например, у хвойных пород обычно равняется 0,5–1,2 м/ч, у дуба – 27–40 м/ч. 3. Факторы, влияющие на поступление воды Внешние факторы 1. Аэрация почвы – обеспеченность кислородом. Для поступления воды нужна энергия, поэтому этот процесс зависит от скорости дыхания корней. Условия, подавляющие дыхание, резко уменьшают поступление воды. Если после сильного дождя в понижении собирается много воды, то растения увядают, т.к. избыток воды вытесняет воздух из почвы и дыхание корней при этом подавляется. Затопление приводит и к увеличению концентрации СО2 в почве. Избыток СО2 повреждает мембраны корневых волосков и тем самым тормозит поступление воды. 2. Температура почвы. На холодных почвах, несмотря на достаточное количество воды, растения испытывают ее недостаток, т.к. при низких температурах угнетается дыхание и нарушается снабжение клеток энергией, необходимой для поглощения солей и, следовательно, поступления воды. В результате у растений формируется структура, характерная для растений жарких сухих мест – ксерофитов: мелкие клетки, много устьиц и проводящих пучков на единице листовой поверхности. Такая структура называется ксероморфной. Состояние, при котором растение испытывает водный дефицит, несмотря на достаточное количество воды в окружающей среде, получило название физиологической засухи. Физиологическая засуха наблюдается не только на холодных, но и на плохо аэрируемых, плохо дренированных, затопляемых водой почвах. Очень сильно реагируют на понижение температуры почвы теплолюбивые растения. Так, у фасоли, томатов, огурцов, тыквы поступление воды прекращается при 5ºС, тогда как деревья тундры поглощают ее и при 0ºС. Это необходимо учитывать при выращивании южных культур в более северных районах. 3. Градиент водного потенциала в системе «почва–растение»: чем меньше концентрация почвенного раствора, тем выше его водный потенциал и тем быстрее вода поступает в корень. Если же почвенный раствор будет иметь более отрицательный осмотический потенциал, то вода не только не будет поступать в корень, но будет выделяться из него в почву. На засоленных почвах относительно низкий водный потенциал почвенного раствора может сильно затруднять поступление воды в корни. 4. Количество воды в почве. Всякое уменьшение влажности почвы снижает поступление воды. Чем меньше воды в почве, тем с большей силой она удерживается и тем меньше ее водный потенциал. Способность почвы удерживать воду зависит от ее состава и свойств. Относительно крупные кристаллы силикатов (песок) связывают воду в значительной степени. Глинистые минералы (алюмосиликаты) и гетерогенные гумусовые вещества, будучи коллоидами, также удерживают значительные количества гидратационной воды. Существует 2 механизма удержания воды в почве: 1) за счет сил, действующих на разделе фаз жидкость–воздух, при этом поверхностное натяжение уравновешивает силы, способствующие удалению воды; 2) благодаря силам, действующим на поверхностях раздела жидкой и твердой фаз. Существуют различные термины, применяемые для обозначения доступности почвенной влаги. Полевая влагоемкость – характеризует максимальный размер запаса почвенной влаги, который может быть использован для роста растения. Влажность устойчивого завядания – показатель минимальных запасов почвенной влаги, при котором растения остаются увядшими до тех пор, пока в почву не поступит вода. Влажность устойчивого завядания представляет собой нижнюю границу того интервала влажности, в котором возможен рост растений. Доступная для растений почвенная влага – количество воды, которое накапливается в почве от уровня влажности устойчивого завядания до полевой влагоемкости. Легкодоступная для растений влага удерживается в почве силой до 0,5 МПа, среднедоступная – до 1–1,2 МПа, труднодоступная – до 2,5–3,0 МПа. Определение доступной почвенной влаги имеет большое практическое значение для выяснения сельскохозяйственной ценности почв, т.к. известно, что в умеренной зоне за вегетационный период растения испаряют больше воды, чем ее выпадает за этот период в виде атмосферных осадков. При этом растения используют накопленную с весны доступную почвенную влагу. По мере иссушения почвы скорость движения воды в ней значительно замедляется. Виды почвенной влаги С физиологической точки зрения выделяют следующие формы почвенной влаги. Различающиеся по степени доступности их для растений. 1. Гравитационная вода – заполняет крупные промежутки между частицами почвы. Хорошо доступна растениям. Водный потенциал этой формы воды составляет -0,1 бар. Однако она легко стекает в нижние горизонты под действием силы тяжести. Бывает в почве лишь после дождей. 2. Капиллярная вода – заполняет капиллярные поры в почве. Эта вода также хорошо доступна растениям. Она удерживается в капиллярах силами поверхностного натяжения и поэтому не стекает вниз, а даже поднимается вверх от грунтовых вод (ψВ не более -1 бар). 3. Пленочная вода – окружает коллоидные частицы почвы. Чем ближе к коллоидным частицам располагаются молекулы воды, тем с большей силой они удерживаются и поэтому менее доступны для растений. 4. Гигроскопическая вода – адсорбируется сухой почвой при помещении ее в атмосферу с 95% относительной влажности воздуха. Этот тонкий слой молекул воды удерживается с такой силой, что их ψВ = -1000 бар. Эта форма воды полностью недоступна для растений. Внутренние факторы Поступление воды зависит от роста самого растения. В процессе роста корень движется к воде, в результате ветвления увеличивается количество зон интенсивного поступления воды в корни. С другой стороны, быстрый рост побегов требует большого количества воды. С торможением роста поступление воды уменьшается, поэтому к концу вегетационного периода, вскоре после образования плодов, изучать поступление воды по количеству выделяемой пасоки не удается. Рост корня обычно опережает рост надземных органов. Это связано с тем, что корень должен поставлять воду в образующиеся побеги. При засухе сначала тормозится рост побегов, а рост корней еще длительное время продолжается. В результате в условиях почвенной засухи резко изменяется соотношение размеров корневой системы и надземных органов. Следовательно, изменяя количество воды в почве, можно в некоторой степени управлять ростом. ЛЕКЦИЯ № 13 |