ЛЕКЦИЯ 11_Водный обмен. Водный обмен растений
 Скачать 2.64 Mb. Скачать 2.64 Mb.
|

|
2. Осмотический механизм. В 19 веке движения устьиц связывали с реакцией превращения крахмала в глюкозо-1-фосфат: крахмалфосфорилаза Крахмал + Фн → глюкозо-1-фосфат Реакция обратима и катализируется крахмалфлсфорилазой, широко распространенной в листьях. Крахмал в отличие от глюкозо-1-фосфата не растворяется в воде, поэтому его концентрация не влияет на величину водного потенциала замыкающих клеток. Если крахмал превращается в глюкозо-1-фосфат, то водный потенциал уменьшается, вода поступает в замыкающие клетки – устьице открывается. Если, наоборот, глюкозо-1-фосфат превращается в крахмал, то водный потенциал увеличивается, и вода уходит из замыкающих клеток, устьице закрывается. 3. Гидродинамический механизм. Ширина устьичной щели зависит от тургора не только в замыкающих клетках, но и в примыкающих. Если в примыкающих много воды (сразу после дождя), то они давят на замыкающие клетки, и устьице не может открыться – это гидропассивное закрывание. Гидропассивное открывание устьиц происходит при ослаблении этого сдавливания в условиях слабого дефицита воды. В условиях засухи первыми теряют воду примыкающие клетки. их объем уменьшается. В результате их давление на замыкающие клетки исчезает и устьице открывается. Однако затем оно все-таки закрывается, т.к. теряют тургор и замыкающие клетки – это гидроактивное закрывание. Гидродинамический механизм работает, когда нарушается поступление воды в листовую пластинку. Если корни переносят в питательный раствор с пониженной температурой или перерезают черешок листа, уже через несколько секунд увеличивается ширина устьичной щели. 4. Фотосинтетический механизм. В отличие от других клеток эпидермы замыкающие клетки устьиц содержат хлоропласты. На свету при хорошем водоснабжении устьица открываются тем шире, чем больше интенсивность освещения, причем действующим фактором является синий свет. Усиление синтеза углеводов (при фотосинтезе) в замыкающих клетках увеличивает их водный потенциал (сосущую силу) и вызывает поглощение воды, что способствует открыванию устьиц. Использование разных механизмов для выполнения одной функции – один из главных принципов организации и функционирования клетки – принцип надежности: если один механизм не может работать, то работает другой – и функция продолжает осуществляться. В регуляции функционирования устьиц взаимодействуют прямые и обратные связи. Одна связь предотвращает недостаток СО2, который может быть вызван фотосинтезом. Когда межклеточная концентрация СО2 снижается до уровня, недостаточного для фотосинтеза – это служит сигналом обратной связи и устьица открываются для обмена СО2 с внешней средой. Другой тип связи реагирует на содержание воды в листьях: при снижении содержания воды в тканях устьица закрываются. В результате деятельности этих 2-х типов обратных связей наблюдаются небольшие осцилляции отверстости устьиц (т.е. устьице открывается–закрывается, опять открывается и закрывается). 2. Внеустьичная регуляция транспирации – это прежде всего уменьшение испарения воды в межклетники, которое осуществляется с помощью нескольких механизмов: 1. Механизм начинающегося подсушивания – связан с обезвоживанием стенок клеток мезофилла, с поверхности которых идет испарение. Если в лист приходит меньше воды, то клеточные стенки хлоренхимы начинают подсыхать. Если воды мало, водные мениски в капиллярах между фибриллами целлюлозы становятся вогнутыми, что увеличивает силы поверхностного натяжения и испарение идет медленнее. 2. Способность цитоплазмы связывать воду. На существование этого механизма указывает тот факт, что при одной и той же степени открытости устьиц интенсивность транспирации может сильно меняться. Это связывают с превращением крахмала в глюкозо-1-фосфат в замыкающих клетках и клетках хлоренхимы. В результате происходит уменьшение водного потенциала, а значит и транспирации. 3. Изменение толщины кутикулы, покрывающей листья. Увеличение ее толщины уменьшает транспирацию с поверхности листьев. В кутикуле могут образовываться трещины, которые увеличивают кутикулярную транспирацию. Кроме того, кутикула обладает интересными свойствами: при подсыхании наружных стенок клеток эпидермы слои кутикулы плотнее придвигаются друг к другу и испарение воды уменьшается. При увеличении оводненности эпидермы кутикулы набухает, разрыхляется и кутикулярная транспирация увеличивается. По ночам при более сильном набухании кутикулы кутикулярная транспирация идет быстрее, чем днем. Эти механизмы уменьшения интенсивности транспирации выгодны тем, что не влияют на поглощение СО2. У некоторых растений существуют свои специфические механизмы внеустьичной регуляции. 4. Свертывание листовых пластинок в трубку вдоль срединной жилки – наблюдается у некоторых злаков, когда транспирация превышает поступление воды. Свертывание уменьшает интенсивность транспирации на 10–20%. У банана при сильном водном дефиците обе половинки листа складываются (рис. 3). В результате между ними повышается относительная влажность воздуха и транспирация снижается.  Рис. 3. Сворачивание листьев банана при засухе Некоторые исследователи считают, что по величине угла между половинками листа можно судить о степени водного дефицита. 5. Потеря листьев – при недостатке воды. Например, у сахарного тростника в условиях засухи на побеге может остаться только 1 лист. Листопад у деревьев умеренной зоны – это приспособление к зимней засухе; в тропиках и пустынях – к началу сухого сезона. Связь между устьичной и внеустьичной регуляцией Когда устьица только начинают открываться интенсивность транспирации растет почти линейною. Однако, когда устьица широко открыты, дальнейшее увеличение щели не влияет заметно на интенсивность транспирации. Таким образом, при максимально открытых устьицах скорость транспирации регулирует только внеустьичный механизм. При закрытых устьицах уменьшение кутикулярной транспирации происходит тоже за счет внеустьичной регуляции. Участие разных механизмов в регуляции транспирации у растений разных видов неодинаково. У оливковых деревьев главным образом в регуляции играет устьичный механизм, у хлопчатника преобладает внеустьичный. У растений умеренной зоны внеустьичное регулирование выражено меньше: без закрывания устьиц транспирация может быть уменьшена на 30%. Таким образом, наземным растениям нужно решать сложную задачу: – с одной стороны, они должны обладать большой поверхностью, чтобы поглощать достаточное количество света и СО2; – с другой – по мере увеличения поверхности, растет и расход воды. Эту проблему растение решает разными способами. Во-первых, увеличивается поступление воды за счет роста корня и развития очень большой поглощающей воду поверхности. Во-вторых, трата воды сокращается благодаря кутикуле, волоскам, покрывающим листовые пластинки, и другими приспособлениями. 4. Показатели транспирации Транспирация характеризуется следующими показателями: интенсивностью, продуктивностью, транспирационным коэффициентом и экономностью. 1. Интенсивность, или скорость транспирации (ИТ) – это количество граммов воды, испаренной с 1 м2 поверхности листьев за час. Ее величина зависит от температуры, влажности воздуха и почвы, освещенности и других факторов. У большинства сельскохозяйственных растений ИТ колеблется от 15 до 250 г воды/м2·ч, а ночью может снижаться до 7–20 г воды/м2·ч и даже больше. Если считать, что в дневные часы средняя ИТ у кутикулярных растений 200 г воды/м2·ч, что они транспирируют 10 ч в сутки и что общая площадь листьев на 1 га пашни равняется 5 га, то получается, что растения расходуют 100 т воды/га. Высокая ИТ вызвана тем, что атмосфера имеет очень низкий водный потенциал. Его величина зависит от влажности воздуха. При относительной влажности воздуха 100% водный потенциал равен 0, при уменьшении влажности уменьшается и водный потенциал. Транспирация уменьшает опасность перегрева тканей, т.к. избыточное тепло тратится на превращение воды из жидкости в пар. 2. Транспирационный коэффициент (ТК) – это количество граммов воды. Израсходованной растением при накоплении 1 г сухого вещества. Средняя величина ТК коэффициента варьирует у разных растений от 100 до 1000 г воды/г сухого вещества, чаще всего она равняется 300–500. 3. Продуктивность транспирации (ПТ) – это количество граммов сухого вещества, накопленного в растении при испарении 1000 г воды. Величина ПТ варьирует от 1 до 8 г сухого вещества на 1 л воды. В умеренной зоне для большинства сельскохозяйственных культур она равняется 3. 4. Относительная транспирация (ОТ) – отношение воды, испаряемой листом, к воде, испаряемой со свободной водной поверхности той же площади за один и тот же промежуток времени. 5. Экономность транспирации (ЭТ) – количество испаряемой воды (в мг) на единицу (в кг) воды, содержащейся в растении. Тонколистные растения расходуют за час 39–119%, тогда как растения с мясистыми листьями – 8–20% от общего количества содержащейся в них воды. 5. Влияние внешних и внутренних факторов на движения устьиц Движения устьиц зависят от изменения внешних условий и состояния клеток, органов, организма. 1. Вода – главный фактор, регулирующий устьичные движения, т.к. она влияет на тургорное давление в замыкающих клетках. У какао при дефиците влажности листьев всего 4,5% устьица закрываются через 6 минут. 2. Другим очень важным фактором является концентрация СО2. При ее снижении в межклетниках устьица открываются. Влияние концентрации СО2 на степень открытости устьиц хорошо видно у суккулентов. У многих из этих растений устьица ночью открыты, а днем – закрыты. Ночью в листьях этих растений идет карбоксилирование ФЕП и образование малата. В результате концентрация СО2 снижается и устьица открываются. Днем устьица закрываются, т.к. происходит декарбоксилирование малата и концентрация СО2 увеличивается. 3. Свет косвенно влияет на устьичные движения через участие в фотосинтезе. При освещении в замыкающих клетках идет фотосинтез и устьица открываются. На свету при хорошем водоснабжении устьица открываются тем шире, чем больше освещенность. 4. Температура влияет прежде всего на скорость открывания устьиц: – при температуре ниже 5ºС устьица открываются очень медленно и неполностью; – при температуре 0–5ºС (отрицательные температуры) – они остаются закрытыми; – при температурах выше оптимальных – устьица открываются неполностью; – при 40–45ºС у некоторых видов устьица открываются максимально. Влияние температуры на движения устьиц опосредованно: через ее влияние на скорость фотосинтеза и на скорость дыхания. При температуре выше 30ºС интенсивность дыхания выше интенсивности фотосинтеза. В результате в межклетниках накапливается СО2, и устьица закрываются. При температурах, оптимальных для фотосинтеза, в межклетниках мало СО2 и устьица открыты. Итак, малая интенсивность света, недостаток воды, высокая температура, вызывающие снижение фотосинтеза и повышение интенсивности дыхания приводят к закрыванию устьиц. Высокая влажность корнеобитаемого слоя, снабжение калием способствует открыванию устьиц. Так как на растение обычно действует не один фактор, а несколько и совпадение условий, вызывающих открывание устьиц, случается редко, то они открыты средне. Полное закрытие устьиц вызывает только очень неблагоприятные условия, которые тоже встречаются редко. Из внутренних факторов на состояние устьиц влияют: 1. Содержание воды в тканях растения, т.е. соотношение поступления воды и ее испарение. 2. Гормоны: абсцизовая кислота (АБК), гиббереллины, цитокинины. АБК вызывает быстрое закрывание устьиц. Водный дефицит стимулирует образование АБК: ее концентрация увеличивается со скоростью 0,15 мкг/г сырой массы в час. В плазмалемме замыкающих клеток находятся специальные белки-рецепторы абсцизовой кислоты. Соединение АБК с рецептором приводит к изменениям в структуре плазмалеммы. В результате Н+-помпа не работает и устьичная щель закрывается. У кукурузы. Щавеля, сахарной свеклы устьице закрывается через 3–9 мин после обработки АБК (0,1–10 мкг). АБК играет роль эндогенного антитранспиранта. Антитранспирантами называют вещества, снижающие интенсивность транспирации. Под влиянием высоких концентраций АБК увеличивается проницаемость клеток корня для воды. Следовательно, АБК улучшает поступление воды в корни и затрудняет расход воды листьями, что особенно важно в условиях засухи. 3. Состояние устьиц изменяется в течение суток. У большинства растений во влажные дни устьица открываются на рассвете, максимум открытия – в утренние часы, к полудню щель устьиц начинает уменьшается и закрывается полностью перед заходом солнца. В пасмурную погоду у растений умеренной зоны устьица открыты не так широко, как в ясную. 6. Влияние внешних и внутренних факторов на транспирацию Как физический фактор процесс испарения воды транспирация зависит от влажности воздуха, температуры, ветра, величины испаряющей поверхности. 1. Прежде всего на транспирацию влияет влажность воздуха: чем меньше относительная влажность воздуха, тем ниже его водный потенциал и тем быстрее идет транспирация. Однако, при возникновении сильного водного дефицита транспирация может совсем прекратиться, несмотря на то, что сухость воздуха увеличивается. С увеличением влажности воздуха транспирация уменьшается; при большой влажности воздуха происходит только гуттация. В районах с большим количеством осадков и высокой влажностью воздуха обитают гигрофиты (кислица обыкновенная, калужница болотная, лютик весенний, сердечник луговой и другие теневыносливые растения), развивающиеся под пологом леса и на открытых сырых почвах. Высокая влажность воздуха препятствует нормальному ходу транспирации, следовательно, отрицательно влияет на восходящий ток веществ по сосудам, регуляцию температуры растения и устьичные движения. 2. Температура. С увеличением температуры интенсивность транспирации увеличивается. Температура – источник энергии для испарения воды. Интенсивность транспирации 1 г воды/дм2·ч соответствует затрата энергии 0,1 кал/см2·мин. Охлаждающий эффект транспирации значителен при высокой температуре, низкой влажности воздуха и хорошем водоснабжении. Кроме того, температура выполняет еще и регуляторную функцию, влияя на степень открытости устьиц. 3. Скорость ветра не так сильно влияет на транспирацию, как на испарение со свободной поверхности. Вначале при появлении ветра и увеличении его скорости до 0,4–0,8 м/с интенсивность транспирации возрастает, но дальнейшее усиление ветра почти не влияет на этот процесс (рис. 4). 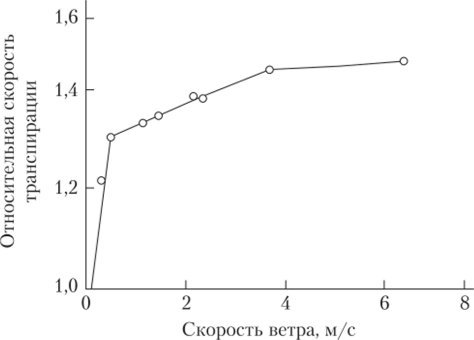 Рис. 4. Влияние скорости ветра на интенсивность транспирации Как физиологический процесс транспирация зависит от света, влажности почвы, удобрений. 1. Свет – главный фактор, регулирующий транспирацию. Интенсивность транспирации у кукурузы в темноте составляет 97, на рассеянном свету – 114, на прямом солнечном свету – 785 мг воды/ дм2·ч. Это связано прежде всего с тем, что зеленые клетки поглощают не только инфракрасные солнечные лучи, но и видимый свет, необходимый для фотосинтеза. Однако лист расходует на фотосинтез только 1–2% поглощенного света (редко 5%), а остальная энергия (до 98%) превращается в тепло и используется на испарение воды. Поэтому влияние света на транспирацию тем сильнее, чем выше концентрация хлорофилла. У зеленых растений даже на рассеянном свету интенсивность транспирации увеличивается на 30–40% по сравнению с темнотой – это прямое действие света на транспирацию. Косвенно свет влияет на открывание–закрывание устьиц. 2. Влажность почвы. С уменьшением влажности почвы уменьшается и транспирация. Чем меньше воды в почве, тем ее меньше в клетках и поэтому растение снижает транспирацию, закрывая устьице. Но если в почве становится очень много воды, то интенсивность транспирации падает, т.к. вода вытесняет О2 из почвы, необходимый для дыхания корней. Нарушение дыхания приводит к плохому поглощению солей и, следовательно, снижению поступления воды, и снижению транспирации. 3. Условия минерального питания. У растений, испытывающих недостаток N, P или K интенсивность транспирации максимальна. Транспирация резко сокращается, если растения получают полное минеральное удобрение. Объясняется это тем, что поглощаемые растением ионы и синтезируемые вещества связывают воду, а испаряется только свободная вода. Следовательно, внесение удобрений – один из методов уменьшения транспирации. Влияние внутренних факторов 1. Содержание воды в листьях. Уменьшение оводненности листьев уменьшает транспирацию. |