ЛЕКЦИЯ 11_Водный обмен. Водный обмен растений
 Скачать 2.64 Mb. Скачать 2.64 Mb.
|

|
ТЕМА: ТРАНСПОРТ ВОДЫ ПО РАСТЕНИЮ 1. Пути и механизм транспорта. 2. Транспирация: значение, этапы, виды. 3. Регуляция транспирации. 4. Показатели транспирации. 5. Влияние внешних и внутренних факторов на движения устьиц. 6. Влияние внешних и внутренних факторов на транспирацию. 7. Суточные колебания транспирации. 8. Способы снижения транспирации. 9. Классификация растений по их водному обмену. 1. Пути и механизм транспорта Транспорт веществ в тканях одного органа называется ближним, а между органами – дальним. Ближний транспорт идет по неспециализированным тканям, для дальнего есть специальные ткани – проводящие. Движение воды осуществляется в 2-х направления: 1. Восходящий ток – вверх по растению: из корней в листья. Основная масса воды транспортируется. 2. Нисходящий ток – вниз. Небольшая часть воды идет. Восходящий ток идет по сосудам (90%) и ситовидным трубкам ксилемы, а нисходящий – только по ситовидным трубкам флоэмы. Лучше изучен восходящий транспорт. Путь, который проходит вода в растении от корневого волоска до испаряющей клетки листа, распадается на 2 части, отличающиеся по длине, строению и физиологически: 1. Первая часть состоит из живых клеток и имеет ничтожные размеры (мм или доли мм). Делится на 2 участка: а) в корне – от корневых волосков до тканей центрального цилиндра; б) в листе – от сосудов, входящих в состав проводящего пучка, до хлоренхимы, испаряющей воду в межклетники. 2. Вторая часть пути – это сосуды, представляющие собой мертвые трубки. У травянистых растений их длина составляет несколько см, а у деревьев – достигает нескольких или десятки метров. От поверхности корня через клетки коры, эндодерму и перицикл вода должна пройти до сосудов ксилемы. Через клетки коры возможны 3 пути транспорта воды и минеральных веществ: 1) через цитоплазму по плазмодесмам (симпластный транспорт); 2) по клеточным стенкам (апопластный транспорт); 3) через вакуоли (трансвакуолярный транспорт), выделяют некоторые авторы. Поскольку сопротивление клеточных стенок для воды значительно ниже, чем у цитоплазмы, поэтому более быстрый транспорт воды идет по апопласту. Транспорт воды по апопласту возможен лишь до эндодермы, т.к. находящиеся в ее клеточных стенках пояски Каспари не пропускают воду. На этом апопластный путь прерывается, и вода переходит на более медленный симпластный путь. Апопласт в корне разделен на 2 части: 1. Первая часть расположена снаружи от эндодермы и включает в себя эпиблему и первичную кору; 2. Вторая – по другую сторону эндодермы и представлена центральным цилиндром. При достаточном водоснабжении вода в коре корня транспортируется главным образом по апопласту, т.к. здесь она встречает меньшее сопротивление, и лишь частично по симпласту. Если клетки корня испытывают недостаток воды, то она будет передвигаться в основном по симпласту. Необходимость переключения транспорта воды с апопласта на симпласт имеет большое значение, т.к. движение по живой цитоплазме клетка может регулировать в отличие от передвижения по мертвым клеточным стенкам. Дальнейший транспорт воды идет по проводящим тканям корня, стебля и листа. Восходящий ток воды идет по сосудам ксилемы, в которых она встречает незначительное сопротивление. Живые клетки оказывают большее сопротивление движению воды, поэтому по ним передвигается лишь 1–10% ее общего количества. Из проводящих тканей стебля вода попадает в проводящие пучки листа. Чем больше проводящих пучков на единице листовой поверхности, тем сильнее они ветвятся, тем меньшее сопротивление встречает вода при своем движении в клетках мезофилла. Иногда мелких проводящих пучков так много, что они проводят воду почти к каждой клетке. Здесь вода из сосуда переходит в клеточные стенки, откуда испаряется в межклетники, следовательно, в листовой пластинке транспорт воды идет в основном по апопласту. Механизм транспорта воды Движущей силой восходящего тока воды в проводящих элементах ксилемы является градиент водного потенциала через растение от почвы до атмосферы. Он поддерживается 2-мя основными механизмами: 1) корневым давлением, или нижним концевым двигателем; 2) транспирацией, или верхним концевым двигателем. 1. Поглощенные клетками эпиблемы вещества не остаются в них. Они транспортируются в соседние клетки первичной коры. А затем поступают в проводящие ткани центрального цилиндра, главным образом, в сосуды. Живые клетки паренхимы центрального цилиндра активно выделяют растворимые органические и минеральные вещества в сосуды. Увеличение концентрации веществ в сосудах понижает их водный потенциал, поэтому вода движется в них через клетки корня. Для активного выделения солей должны работать мембранные транспортные белки, требующие затраты АТФ. Поэтому передвижение воды в корневой системе зависит от дыхания корней, температуры и аэрации почвы, действия дыхательных ядов. Некоторые авторы считают, что живые клетки паренхимы центрального цилиндра активно выделяют в сосуды не только вещества, но и волу с помощью сократительных белков цитоплазмы. По их гипотезе ближний транспорт воды в корне состоит из 2-х последовательных фаз: 1) поступления воды в клетку 2) выделения (выталкивания) ее по направлению к сосудам. Поступление и выталкивание воды происходит в результате сокращения белков и расслабления сократительных белков, которое приводит к изменению объема паренхимных клеток первичной коры и центрального цилиндра. Максимуму выделения воды соответствует сокращение белков и уменьшение объема клетки, а минимуму – расслабление сократительных белков и увеличение ее объема. Сократительные – актиномиозиновые – белки были выделены из клеток огурца и подсолнечника. Выделение воды приводит к уменьшению водного потенциала клетки, создавая тем самым условиям для поступления в нее следующей порции воды, в результате транспорт происходит ритмично. 2. Вторая причина возникновения градиента водного потенциала в растении – испарение воды побегами или транспирация. Чем интенсивнее клетки листьев испаряют воду, тем быстрее она будет поступать в них из сосудов, а в сосуды – из клеток корней и быстрее подниматься по растению. Вода может подниматься вверх на десятки метров благодаря силам сцепления (явление когезии), которые возникают между ее молекулами за счет водородных связей. Теория сцепления была предложена Г. Диксоном (Англия, 1895) и Е.Ф. Вотчалом (Россия, 1897). По этой теории молекулы воды, соединяясь друг с другом образуют в растении сплошные водяные нити, проходящие от клеток эпиблемы корня до испаряющих клеток паренхимы листовой пластинки. Поэтому испарение одной молекулы воды заставляет двигаться вверх всю водяную нить. Силы сцепления достигают величины 3,0 МПа. Бòльшая часть водного тока испаряется в атмосферу в процессе транспирации, а меньшая (около 0,2%) используется: 1) на метаболические реакции, входя в состав синтезируемых растением органических соединений; 2) на поддержание тургора клеток; 3) в транспорте органических веществ по флоэме от листьев к корням. Значение восходящего тока для растений 1. Транспорт и накопление в надземных органах минеральных веществ и химических соединений, поступающих их корней. Поэтому в растениеводстве для получения максимальных урожаев необходимо знать соотношение между продукцией фотосинтеза и расходованием воды у выращиваемых растений с целью их регуляции. 2. Поддержание нормального водоснабжения всех клеток и соответственно тургора. Поэтому в засушливых районах необходимо развивать орошаемое земледелие. В условиях высокой сухости почвы большую роль в поддержании водного режима растений может играть их опрыскивание (создание искусственной росы). Нисходящий ток Направленный вниз флоэмный ток растворов формируется в клетках мезофилла листьев, где часть воды, доставленной ксилемным током, из клеточных стенок мезофилла переходит во флоэмные окончания. Вода из клеток листа и сосудов ксилемы поступает во флоэму по осмотическому градиенту, который возникает вследствие накопления в клетках флоэмы сахаров и других органических соединений, образуемых в процессе фотосинтеза. Нисходящий флоэмный ток доставляет органические соединения тканями корня, где они используются в метаболизме. В корне элементы флоэмы, как и в листе, располагаются вблизи элементов ксилемы и вода вновь по осмотическому градиенту поступает в ксилему и движется вверх с восходящим током. Таким образом происходит обмен воды в проводящей системе корня и листьев. Транспорт воды из корня в лист по мертвым сосудам, оказывающим минимальное сопротивление водному току, – одно из величайших изобретений природы. Для транспорта воды даже на небольшие расстояния по живым клеткам понадобилась бы разница в водных потенциалах в 100 МПа, в действительности же этого нет. Выход в процессе эволюции растений на сушу, выносящих свою крону на несколько метров (иногда десятки метров) над поверхностью земли, стал возможен благодаря образованию проводящих тканей. Значение этого приспособления подчеркивается и самим названием высших растений – сосудистые. 2. Транспирация: значение, этапы, виды Листья выделяют воду в результате транспирации и гуттации. Про гуттацию было рассказано в предыдущей лекции. Транспирация – это физиологический процесс испарения воды надземными органами растения. Транспирация возможна лишь тогда, когда окружающий побеги воздух не насыщен водой. В противном случае происходит гуттация. Транспирация – это основной путь расходования воды растением. Значение транспирации 1. Своеобразный терморегулятор: большая часть солнечной энергии, поглощаемой листьями, расходуется на превращение воды в пар, поэтому температура листьев мало изменяется даже в очень жаркие дни. 2. Поддерживает устьица открытыми для нормального газообмена. 3. Транспирация работает как верхний концевой двигатель и помогает веществам и воде подниматься вверх по растению. Лист как орган транспирации Главный орган транспирации – лист, хотя могут транспирировать и стебли. Общий план строения листа известен из курса ботаники. Рассмотрим те особенности анатомии, которые влияют на транспирацию. Сверху и снизу лист покрыт эпидермой, которая состоит из одного слоя клеток, тесно примыкающих друг к другу. Наружные стенки этих клеток покрыты кутикулой. Входящие в ее состав вещества: кутин, воск – гидрофобны. Это затрудняет испарение воды и позволяет эпидерме поддерживать водный гомеостаз листа. Из стебля вода движется в лист через черешок или листовое влагалище, а затем по листовым жилкам. Количество проводящих элементов в каждой жилке последовательно уменьшается по мере их ветвления. Самые мелкие жилки состоят из единичных трахеид. Система распределения настолько эффективна, что редко клетки листа отделены от сосудистых элементов более чем двумя другими клетками. Жидкая вода транспортируется к испаряющим поверхностям преимущественно по клеточным стенкам. Как и в корне это обусловлено тем, что здесь она встречает меньшее сопротивление, чем на пути от клетки к клетке через протопласты и вакуоли. В межклетниках воздух насыщен водой наполовину, а водный потенциал уравновешен с водным потенциалом окружающих клеток, где он редко бывает 2 МПа. Молекулы воды покидают растение перемещаясь (как и внутри растения) в направлении более низкого водного потенциала, т.е. из тканей наружу через устьица, которые расположены в эпидерме. Число устьиц и их расположение в листе различны. Они могут находиться на обоих сторонах листа (у травянистых растений), или только на нижней (у деревьев). В среднем число устьиц колеблется от 50 до 500 на 1 мм2. Длина устьичной щели 20–30, а ширина 4–6 мкм. Транспирация с поверхности листа через устьица идет почти с такой же скоростью, как и с поверхности чистой воды (50–70%). Это объясняется законом Стефана: испарение с малых поверхностей (площадь устьичной щели) пропорциональна не их площади, а диаметру. Устьице – щель между двумя замыкающими клетками, в которых имеются хлоропласты, отсутствующие в других клетках эпидермы (рис. 1). Особенности строения замыкающих клеток – неодинаковая толщина их стенок: стенки, примыкающие к устьичной щели утолщены, а противоположные им – тонкие и эластичные. При увеличении тургорного давления в замыкающих клетках тонкие части стенки растягиваются и выпячиваются, а толстые, обращенные к щели – становятся вогнутыми – щель расширяется и открывается. В замыкающих клетках идет активный фотосинтез благодаря наличию хлоропластов. здесь находится больше осмотически активных веществ, отчего концентрация раствора в ней выше, чем в прилегающей. 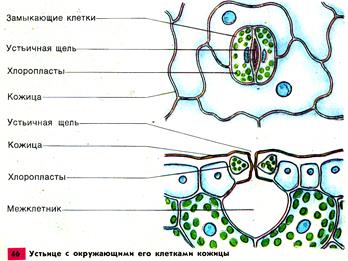 Рис. 1 Строение устьица Между нижней и верхней эпидермой находится мезофилл с системой межклетников и проводящими пучками (рис. 2). Межклетники увеличивают внутреннюю испаряющую поверхность листа в 7–10 раз и соединяются с внешней средой через устьица. Поверхность всех клеток мезофилла в 10–30 раз больше поверхности листовой пластинки, что значительно увеличивает площадь испарения. Вода в межклетниках испаряется со всех открытых участков клеток мезофилла. На поверхности эпидермы часто развиты волоски, чешуйки, которые также влияют на водный режим листа, т.к. сильно снижают скорость движения воздуха над его поверхностью и рассеивают свет, что снижает потери воды за счет транспирации. Устьица играют важную роль в газообмене между листом и атмосферой. Это основной проводящий путь для водяного пара, СО2 и О2. 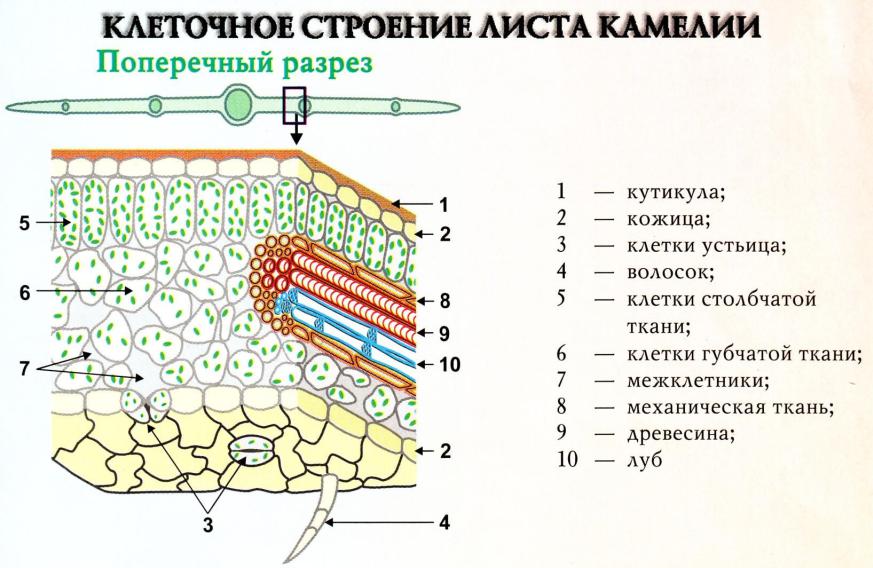 Рис. 2. Клеточное строение листа камелии Транспирация слагается из 3-х этапов: 1. Передвижение воды из листовых жилок в поверхностные слои стенок мезофилла. 2. Испарение воды из клеточных стенок в межклетные пространства и подустьичные полости. 3. Последующая диффузия воды в окружающую атмосферу через устьица (устьичная транспирация) или испарение воды из клеточных стенок эпидермиса в атмосферу путем кутикулярной транспирации. Виды транспирации: 1) устьичная – испарение воды через устьица; 2) кутикулярная – испарение воды через всю поверхность листовой пластинки; 3) перидермальная – испарение с поверхностей ветвей и стволов древесных растений через чечевички и окружающие их слои пробки. Кутикулярная транспирация При открытых устьицах потери водяного пара через кутикулу листа обычно незначительны по сравнению с общей транспирацией. Но если устьица закрыты (во время засухи) кутикулярная транспирация приобретает важное значение в водном режиме растений многих видов. Интенсивность кутикулярной транспирации сильно варьирует у разных видов: от незначительных потерь до 50% от общей транспирации. У молодых листьев кутикулярная транспирация составляет 30–50% общей интенсивности транспирации; у взрослых – она в 10 раз слабее. У кактусов кутикулярная транспирация практически отсутствует. Транспирировать могут не только листья, но и стебли и некоторые другие органы растений в незначительных количествах (до 10%). Например, репродуктивные органы: корзинки подсолнечника, коробочки мака и плоды перца транспирируют сильнее, чем листья данных растений в тех же условиях. Кутикулярная транспирация регулируется главным образом толщиной слоя кутикулы. Виды магнолий и хвойных, обладающих толстыми слоями кутина в кутикуле, теряют очень мало воды. И наоборот, виды с тонким слоем кутикулы продолжают терять воду после того, как устьица закроются, и поэтому гораздо сильнее страдают от засухи. 3. Регуляция транспирации Различают 2 типа регуляции транспирации: устьичный и внеустьичный. 1. Устьичная регуляция осуществляется с помощью открывания и закрывания устьиц. Закрывание устьиц наполовину мало влияет на интенсивность транспирации. Что вытекает из закона Стефана. Полное их закрывание сокращает транспирацию примерно на 90%. Механизмы устьичных движений Состояние устьица зависит от количества воды в замыкающих клетках. Устьице открывается, если в замыкающие клетки поступает вода, что приводит к увеличению тургорного давления в этих клетках и к изменению их формы. Если из замыкающих клеток вода уходит, тургорное давление в них падает, форма клеток изменяется и устьице закрывается. Существует несколько механизмов, влияющих на изменение тургорного давления в замыкающих клетках. 1. Калиевый насос. Тургорное давление зависит от перераспределения ионов калия между замыкающими и примыкающими клетками эпидермы. В ответ на изменение внешних и внутренних условий на плазмалемме замыкающих клеток начинает работать Н+-помпа, откачивающая протоны из этих клеток в примыкающие. Это активный процесс, для которого поставщиком АТФ является дыхание. Вместо протонов в замыкающие клетки поступают ионы К+. в результате в этих клетках понижается водный потенциал и в них поступает вода. Доказательством выхода протонов из замыкающих клеток служит повышение величины рН при поступлении в них ионов К+. Источником протонов могут быть органические кислоты клеточного сока. Иногда протоны не выходят из замыкающих клеток, тогда вслед за К+ устремляются ионы Cl-. При выделении ионов калия из замыкающих клеток происходит повышение водного потенциала в замыкающих клетках, вода выделяется в примыкающие клетки и устьица закрывются. |