Полушин Ю.С Руководство. Руководство по анестезиологии и реаниматологии под редакцией профессора Ю. С. Полушина
 Скачать 4.32 Mb. Скачать 4.32 Mb.
|

3.2. ДыханиеДыхание - процесс доставки кислорода (О2) к клеткам организма и использование его в биологическом окислении органических веществ с образованием воды и углекислого газа (СО2), который выводится в атмосферу. Эффективный газообмен возможен при интеграции и координации функций различных органов, которые в совокупности образуют систему дыхания. Последняя включает следующие подсистемы: "внешнее дыхание" (газообмен в легких, через кожу и слизистые оболочки), транспорт газов кровью (дыхательную функцию крови и сердечно-сосудистой системы) и тканевое дыхание (процесс биологического окисления в клетке, сопровождающийся поглощением тканями О2 и выделением СО2). Газообмен в легких (или «легочное дыхание») обеспечивается легкими с дыхательными путями и капиллярным кровотоком, грудной клеткой с дыхательными мышцами, аппаратом управления. С помощью легочного дыхания осуществляется обмен О2 и СО2 между атмосферным воздухом и артериальной кровью. Газообменная функция легких - одна из важнейших. Ее реализация определяется тремя основными механизмами: вентиляцией, кровотоком и диффузией газов. Транспорт газов (перенос О2 из легочных капилляров в капилляры тканей и СО2 в обратном направлении) зависит в основном от работы "насоса" сердечно-сосудистой системы и дыхательной функции крови. Соответственно, нарушения его можно разделить на гемодинамические (при сердечной и сосудистой недостаточности) и гемические (уменьшение количества циркулирующего гемоглобина, ухудшение связывания и отдачи дыхательных газов, нарушение растворимости их в крови). Тканевое дыхание (процесс энергетического обмена) практически во всех клетках человеческого организма происходит аэробным путем, т. е. с использованием кислорода. Окислительное фосфорилирование потребляет более 90 % поступающего в организм кислорода. Оно происходит с участием ферментов (цитохромов) и направлено на синтез АТФ. Основным источником энергии клетки является глюкоза, для метаболизации которой и нужен О2: С6Н12O6 + 6О2 → 6СО2 + 6Н2О + Энергия При окислении одной молекулы глюкозы образуется 38 молекул АТФ. Энергия, аккумулированная в АТФ, используется для работы ионных насосов, мышечного сокращения, синтеза белка или клеточной секреции. Организм не способен создавать запасы АТФ и должен его постоянно синтезировать, а это требует непрерывной доставки метаболических субстратов и кислорода к клеткам. При анаэробном метаболизме, идущем без участия кислорода, образуется всего 2 молекулы АТФ (при превращении пирувата в молочную кислоту). Более того, развивающийся лактат-ацидоз резко ограничивает активность ферментов, участвующих в превращениях. Когда доставка кислорода к тканям улучшается, лактат вновь преобразуется в пируват, и аэробный метаболизм возобновляется. 3.2.1. Внешнее дыханиеСтруктура воздухоносных путей (ВП) и паренхимы легких. Воздухоносные (или дыхательные) пути соответственно их положению в теле подразделяют на верхние и нижние. К верхним ВП относят полость носа, носовую часть глотки, ротовую часть глотки, к нижним ВП – гортань, трахею, бронхи, включая внутрилегочные разветвления бронхов. Увлажнение и фильтрация вдыхаемого воздуха осуществляется в верхних дыхательных путях (в носу, во рту и в глотке). Нижние воздухоносные пути представляют собой систему дихотомически ветвящихся трубок (каждый бронх разветвляется на два меньших бронха). Хотя диаметр каждой дочерней ветви меньше диаметра родительской трубки, от которой она берет начало, общая площадь поперечного сечения каждой последующей генерации ВП возрастает из-за значительного увеличения общего числа ветвей. 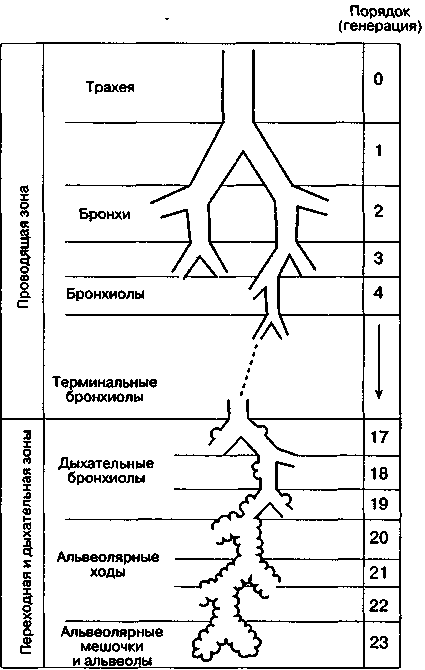 Рис. 3.1. Упрощенная схема воздухоносных путей человека (по Е. R. Weibel, 1963) В легком человека насчитывается в среднем 23 генерации ВП (рис.3.1). Первые 16 называют проводящими, так как они обеспечивают поступление газа к зонам легких, где происходит газообмен. Поскольку в проводящих воздухоносных путях нет альвеол и, следовательно, они не могут участвовать в газообмене, в совокупности образуемую ими емкость называют анатомическим мертвым пространством. Объем его составляет около 150 мл. Проводящие ВП включают бронхи, бронхиолы и терминальные бронхиолы. Последние семь генераций ВП состоят из дыхательных бронхиол, альвеолярных ходов и альвеолярных мешочков. Каждое из этих образований дает начало альвеолам. Дыхательная бронхиола первого порядка (Z=17) и все дистально расположенные от нее газообменивающие ВП образуют легочный ацинус. Вдыхаемый воздух продвигается примерно до конечных бронхиол по механизму объемного потока, однако из-за возрастания общей площади поперечного сечения ВП, вследствие многократных ветвлений, поступательное перемещение газов становится очень незначительным. Главным механизмом вентиляции в дыхательной зоне является диффузия газов. Стенки трахеи и бронхов состоят из трех основных слоев: внутренней слизистой оболочки; гладкомышечного слоя, отделенного от слизистой соединительнотканной подслизистой прослойкой; и внешнего соединительнотканного слоя, содержащего в больших бронхах и трахеи хрящ. Бронхиальный эпителий является псевдослоистым, состоящим из высоких и низких базальных клеток, каждая из которых прикреплена к базальной мембране. При каждом делении элементов трахеобронхиального дерева характер эпителия их слизистой оболочки и подлежащих структур постепенно меняется. Эпителий переходит от реснитчатого столбчатого к кубическому и затем к плоскому альвеолярному. Газообмен может осуществляться только через плоский эпителий, который появляется в дыхательных бронхиолах (бронхи 17-19-го порядка). Эпителиальные клетки ВП несут на своей апикальной поверхности реснички, которые являются важными элементами мукоцилиарной системы. Реснички столбчатого и кубического эпителия синхронно колеблются в направлении носоглотки, продвигая защитный слой слизи, секретируемой бокаловидными клетками, расположенными между реснитчатыми клетками эпителия. Гладкая мускулатура ВП, собранная в непрерывные пучки внутри соединительнотканной подслизистой прослойки, простирается от главных бронхов до дыхательных бронхиол. Мышечные пучки проникают также в газообменные зоны, располагаясь в стенках у входа в альвеолы. Стенки дыхательных путей постепенно теряют хрящевую основу (в бронхиолах) и гладкую мускулатуру. Утрата хрящевой основы приводит к тому, что с уменьшением диаметра проходимость дыхательных путей становится зависимой от радиального растяжения, обусловленного эластическими структурами окружающих тканей. Вследствие этого диаметр мелких дыхательных путей определяется общим объемом легких. Из общей емкости легких (5 л) большая часть (около 3 л) приходится на дыхательную (газобменную) зону. Она включает около 300 млн. альвеол. Поверхность альвеолярно-капиллярного барьера составляет 50-100 м2, а толщина - 0,5 мкм. Эпителий, выстилающий внутреннюю поверхность альвеолы, состоит из двух типов клеток: плоских выстилающих (I тип) и секреторных (II тип) Клетки первого типа занимают до 95% площади альвеолярной поверхности. Клетки второго типа продуцируют и секретируют сурфактант, состоящий из протеинов и фосфолипидов. Он распределяется по альвеолярной поверхности и снижает поверхностное натяжение. Эндотелий капилляров также состоит из слоя плоских выстилающих клеток, располагающихся на эндотелиальной базальной мембране. В зоне альвеол базальные мембраны эпителия и эндотелия спаяны друг с другом, что создает сверхтонкий барьер для обмена газов. В отличие от тесного контакта соседних эпителиальных клеток, образующих непроницаемый барьер, соединения между эндотелиальными клетками довольно слабые. Это позволяет воде и растворенным в ней веществам перемещаться между плазмой и интерстициальным пространством (рис. 3.2). 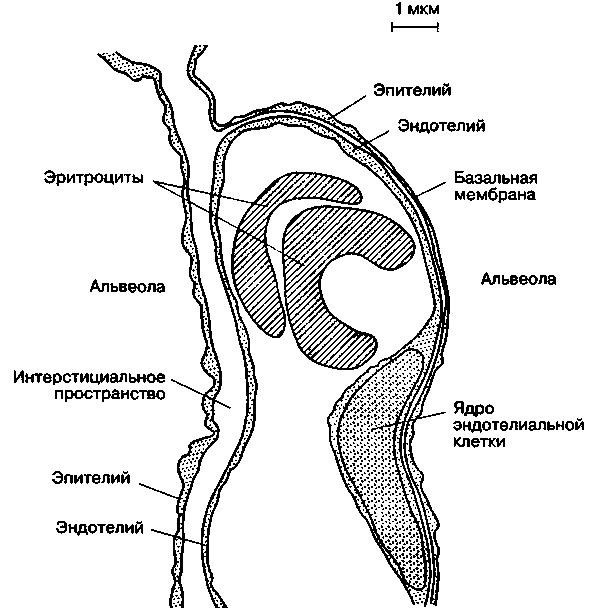 Рис.3.2. Альвеолярно-капиллярная мембрана. Показаны легочное интерстициальное пространство с капилляром, проходящим между двумя альвеолами. Капилляр выпячивается в просвет расположенной справа альвеолы через ее тонкую (газообменную) стенку. Интерстициальное пространство сливается с толстой стенкой левой альвеолы (по J. F. Nunn, 1987) Механика дыхания - область физиологии дыхания, которая рассматривает силы, ответственные за движения потока воздуха внутрь грудной клетки и обратно. Для того чтобы обеспечить поглощение кислорода и выделение двуокиси углерода, свежий воздух должен постоянно доставляться к альвеолам с помощью дыхательного насоса. Понимание механизма действия дыхательного насоса, обеспечивающего поток газа в легкие, требует рассмотрения функции дыхательных мышц, свойств ВП, грудной клетки и легких, которые определяют импеданс системы дыхания, а также причин неравномерности вентиляции и других факторов. При спонтанном дыхании активность дыхательных (инспираторных) мышц необходима для преодоления импеданса дыхательной системы (эластичность × сопротивление). Важнейшей мышцей вдоха является диафрагма. К ней подходят нервы от 3-го, 4-го и 5-го шейных сегментов спинного мозга. При сокращении диафрагмы органы брюшной полости смещаются вниз и вперед, вследствие чего вертикальные размеры грудной полости возрастают. Кроме того, при этом поднимаются и расходятся ребра, приводя к увеличению ее поперечника. При спокойном дыхании диафрагма смещается примерно на 1 см, а при форсированном амплитуда ее движений может достигать 10 см. Поскольку диафрагма имеет куполообразную форму, особое значение приобретает отношение между давлением, напряжением и радиусом ее кривизны. В соответствие с законом Лапласа оно описывается следующим уравнением: Р=2Т/r(3.1), где: Р- давление создаваемое мышцей, Т - напряжение мышцы, r - радиус кривизны. По мере уплощения диафрагмы радиус ее кривизны увеличивается и генерируемое давление снижается. Это явление, например, вместе с укорочением мышцы обусловливает снижение силы диафрагмы у пациентов с хронической обструктивной болезнью легких. При спокойном дыхании диафрагма является единственной активной инспираторной мышцей. При необходимости увеличения вентиляции, например при физической нагрузке или болезненных состояниях, подобных бронхиальной астме, активизируются и другие мышцы. К ним относятся наружные межреберные, лестничные и грудино-ключично-сосцевидные. Две последние группы мышц называются дополнительными дыхательными мышцами. В отличие от вдоха, выдох в нормальных условиях в состоянии покоя происходит пассивно. Эластическая отдача легких и грудной стенки обеспечивает возникновение градиента давления, достаточного для экспираторного потока. При обструкции ВП выдох становится активным процессом, требующим работы экспираторных мышц, включая внутренние межреберные и брюшные (наружную и внутреннюю косою, поперечную брюшную и прямую брюшную). Сокращение инспираторных мышц создает градиент давления между атмосферой и альвеолами, в результате чего возникает поток воздуха. Этот градиент преодолевает: (1) эластическую отдачу дыхательной системы, (2) фрикционное сопротивление ВП воздушному потоку и (3) инерционное сопротивление трахеобронхиального воздушного столба, легких и грудной стенки. Взаимоотношения этих трех элементов выражаются уравнением движения легких: P = (E •V) + (R V ’)+(I +V’’) (3.2), где: P - движущее давление; E - эластичность; V - изменение объема легких; R - сопротивление; V ’- объемная скорость потока воздуха; I - инерционность; V’’ - скорость изменения объемной скорости воздушного потока (ускорение). Таким образом, механические свойства системы дыхания, от которых зависит эластическое и неэластическое сопротивление воздушному потоку, определяют необходимое движущее давление. В норме основная работа дыхательных мышц затрачивается на преодоление эластического сопротивления легких и грудной клетки. Причем, отношение между давлением и изменением объема легких не остается постоянным во всем диапазоне легочных объемов. При их малой величине это отношение может быть выражено как: Р = где: Е - эластичность; Эластичность - есть мера упругости легочной ткани. Чем больше эластичность ткани, тем больше давления требуется приложить для достижения заданного изменения объема легких. Эластическая тяга легких возникает благодаря высокому содержанию в них эластиновых и коллагенновых волокон. Эластин и коллаген находятся в альвеолярных стенках вокруг бронхов и кровеносных сосудов. Возможно, упругость легких обусловлена не столько удлинением этих волокон, сколько изменением их геометрического расположения, как это наблюдается при растяжении нейлоновой ткани: хотя нити сами по себе не изменяют длины, ткань легко растягивается благодаря их особому переплетению. Определенная доля эластической тяги легких обусловлена также действием сил поверхностного натяжения на границе "газ-жидкость" в альвеолах. Поверхностное натяжение - это сила, возникающая на поверхности, разделяющей жидкость и газ. Она обусловлена тем, что межмолекулярное сцепление внутри жидкости гораздо сильнее, чем силы сцепления между молекулами жидкой и газовой фазы. В результате этого площадь поверхности жидкой фазы становится минимальной. Силы поверхностного натяжения в легких взаимодействуют с естественной эластической отдачей, обеспечивая спадение альвеол. Специальное вещество (сурфактант), состоящее из фосфолипидов и протеинов и выстилающее альвеолярную поверхность, снижает внутриальвеолярное поверхностное натяжение. Сурфактант секретируется альвеолярными эпителиальными клетками II типа и выполняет несколько важных физиологических функций. Во-первых, понижая поверхностное натяжение, он увеличивает растяжимость легкого (уменьшает упругость). Тем самым уменьшается совершаемая при вдохе работа. Во-вторых, обеспечивается стабильность альвеол. Давление, создаваемое силами поверхностного натяжения в пузырьке (альвеоле), обратно пропорционально его радиусу, поэтому при одинаковом поверхностном натяжении в мелких пузырьках (альвеолах), оно больше, чем в крупных. Эти силы также подчиняются закону Лапласа, упомянутому ранее (1), с некоторой модификацией: «Т» – поверхностное натяжение, а «r» - радиус пузырька. В отсутствие природного детергента мелкие альвеолы стремились бы перекачать свой воздух в более крупные (рис. 3.3.). Поскольку при изменении диаметра меняется слойная структура сурфактанта, его эффект в отношении снижения сил поверхностного натяжения проявляется тем больше, чем меньше диаметр альвеол. Последнее обстоятельство сглаживает эффект меньшего радиуса кривизны и увеличенного давления. Тем самым предотвращается спадение альвеол и появление ателектазов на выдохе (диаметр альвеол минимален), а также перемещение воздуха из меньших альвеол внутрь больших альвеол (за счет выравнивания сил поверхностного натяжения в альвеолах разного диаметра). 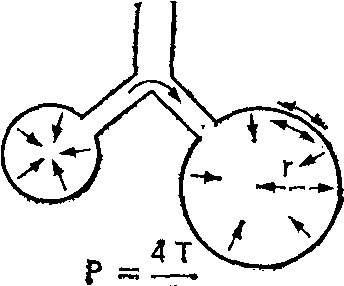 Рис. 3.3. При данном поверхностном натяжении газ из меньшей альвеолы будет перемещаться в больший, поскольку меньший радиус кривизны (r1r2) создает более высокое давление (Р1Р2) в меньшем пузырьке (закон Лапласа) Респираторный дистресс-синдром новорожденных характеризуется дефицитом нормального сурфактанта. У больных детей легкие становятся ригидными, неподатливыми, склонными к коллапсу. Дефицит сурфактанта имеется и при респираторном дистресс-синдроме взрослых, однако, его роль в развитии этого варианта дыхательной недостаточности менее очевидна. Давление, создаваемое эластической паренхимой легкого называется давлением эластической отдачи (Pel). В качестве меры эластической тяги обычно используют растяжимость (С – от англ. complianсе), которая находится в реципрокном отношении к эластичности: С = 1/Е = ΔV/ΔP (3.4), где: С – растяжимость (комплайнс); Е – эластичность (см. 3.3). 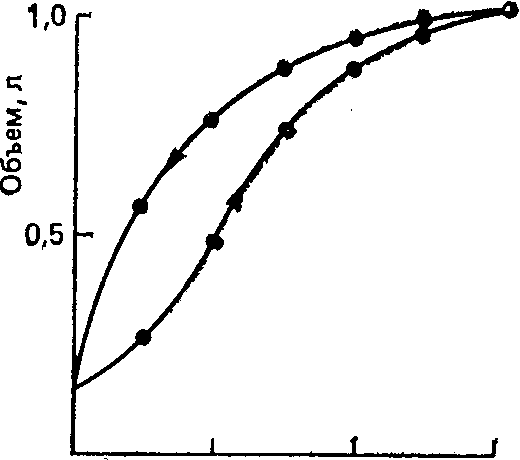 Рис. 3.4.Кривая «объем-давление» Растяжимость (изменение объема на единицу давления) отражается наклоном кривой «объем-давление». Из рисунка 3.4 видно, что такие кривые при раздувании и спадении легких неодинаковы - при одном и том же давлении объем спадающихся легких больше, чем во время их раздувания. Подобные различия между прямым и обратным процессом называются гистерезисом. Кроме того, видно, что кривые не исходят из начала координат. Это указывает на то, что легкое содержит небольшой, но измеримый объем газа даже тогда, когда на него не действует растягивающее давление. Растяжимость обычно измеряется в статических условиях (Сstat), т. е. в состоянии равновесия или, другими словами, в отсутствии движения газа в дыхательных путях. Динамическая растяжимость (Cdyn), которую измеряют на фоне ритмичного дыхания, зависит еще и от сопротивления дыхательных путей. На практике Сdyn измеряется по наклону линии, проведенной между точками начала вдоха и выдоха на кривой «динамическое давление-объем» (рис. 3.5). Рис. 3.5. Кривая «динамическое давление-объем» В физиологических условиях статическая растяжимость легких человека при небольшом давлении (5-10 см Н2О) достигает примерно 200 мл/см вод. ст. При более высоких давлениях (объемах) она, однако, уменьшается. Этому соответствует более пологий участок кривой «давление—объем». Растяжимость легких несколько снижается при альвеолярном отеке и коллапсе, при повышении давления в легочных венах и переполнении легких кровью, при увеличении объема внесосудистой жидкости, наличии воспаления или фиброзе. При эмфиземе легких растяжимость возрастает, как считают, за счет утраты или перестройки эластических составляющих легочной ткани (рис. 3.6). Рис.3.6. Кривые давление-объем у здоровых и больных людей Поскольку изменения давления и объема нелинейны, для оценки упругих свойств легочной ткани часто используют «нормализованную» растяжимость, отнесенную к единице объема легких - |