Полушин Ю.С Руководство. Руководство по анестезиологии и реаниматологии под редакцией профессора Ю. С. Полушина
 Скачать 4.32 Mb. Скачать 4.32 Mb.
|

3.2.2. Транспорт газов кровьюКислород транспортируется артериальной кровью в двух формах: связанный с гемоглобином внутри эритроцита и растворенный в плазме. Эритроцит происходит из недифференцированной костномозговой ткани. При созревании клетка утрачивает ядро, рибосомы и митохондрии. Вследствие этого эритроцит не способен к выполнению таких функций, как клеточное деление, окислительное фосфорилирование и синтез белка. Источником энергии для эритроцита служит преимущественно глюкоза, метаболизируемая в цикле Эмбдена-Миергофа, или гексозомонофосфатном шунте. Наиболее важным внутриклеточным белком для обеспечения транспорта О2 и СО2 является гемоглобин, представляющий собой комплексное соединение железа и порфирина. С одной молекулой гемоглобина связываются максимально четыре молекулы О2. Гемоглобин, полностью загруженный О2, называется оксигемоглобином, а гемоглобин без О2 или присоединивший менее четырех молекул О2 - деоксигенированным гемоглобином. Основной формой транспорта О2 является оксигемоглобин. Каждый грамм гемоглобина может максимально связать 1,34 мл О2. Соответственно, кислородная емкость крови находится в прямой зависимости от содержания гемоглобина: О2 емкость крови = [Hb] 1,34 О2 /гHb/100 мл крови (3.21). У здоровых людей с содержанием гемоглобина 150 г/л кислородная емкость крови составляет 201 мл О2 крови. Кровь содержит незначительное количество кислорода, не связанного с гемоглобином, а растворенного в плазме. Согласно закону Генри, количество растворенного О2 пропорционально давлению О2 и коэффициенту его растворимости. Растворимость О2 в крови очень мала: только 0,0031 мл растворяется в 0,1 л крови на 1 мм рт. ст. Таким образом, при напряжении кислорода 100 мм рт. ст. в 100 мл крови содержится только 0,31 мл растворенного О2. Содержание кислорода в крови (СаО2) - это сумма связанного с гемоглобином и растворенного в плазме О2: СаО2 = [(1,34)[Hb](SaО2)] + [(Pa)(0,0031)] (3.22). Кривая диссоциации гемоглобина. Сродство гемоглобина к кислороду возрастает по мере последовательного связывания молекул О2, что придает кривой диссоциации оксигемоглобина сигмовидную или S-образную форму (рис. 3.14). Верхняя часть кривой (РаО260 мм рт.ст.) плоская. Это указывает на то, что SaО2 и, следовательно, СаО2, остаются относительно постоянными, несмотря на значительные колебания РаО2. Повышение СаО2 или транспорта О2 может быть достигнуто за счет увеличения содержания гемоглобина или растворения в плазме (гипербарическя оксигенация). РаО2, при котором гемоглобин насыщен кислородом на 50% (при 370 рН=7,4), известно как Р50. Эта общепринятая мера сродства гемоглобина к кислороду. Р50 крови человека составляет 26,6 мм рт. ст. Однако оно может изменяться при различных метаболических и фармакологических условиях, воздействующих на процесс связывания кислорода гемоглобином. К ним относят следующие факторы: концентрацию ионов водорода, напряжение углекислого газа, температуру, концентрацию 2,3-дифосфоглицерата (2,3-ДФГ) и др. 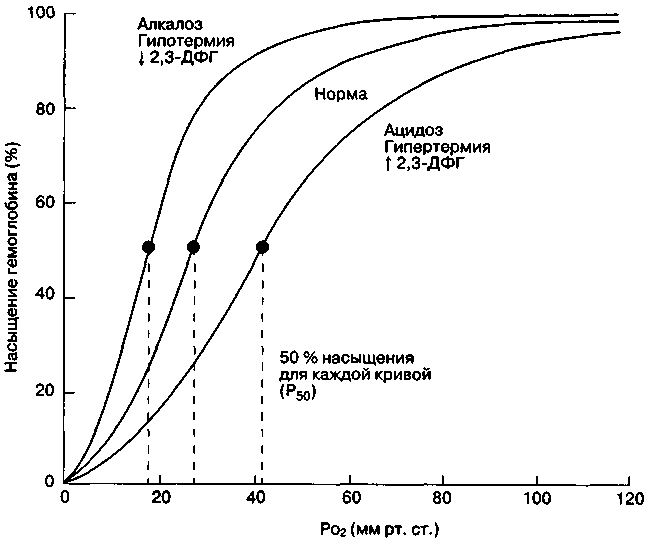 Рис. 3.14. Сдвиги кривой диссоциации оксигемоглобина при изменениях рН, температуры тела и концентрации 2,3-дифосфоглицерата (2,3-ДФГ) в эритроцитах Изменение сродства гемоглобина к кислороду, обусловленное колебаниями внутриклеточной концентрации водородных ионов, называется эффектом Бора. Снижение рН сдвигает кривую вправо, повышение рН - влево. Форма кривой диссоциации оксигемоглобина такова, что этот эффект более выражен в венозной крови, чем в артериальной. Данный феномен облегчает освобождение кислорода в тканях, практически не сказываясь на потреблении кислорода (в отсутствии тяжелой гипоксии). Двуокись углерода оказывает двоякое действие на кривую диссоциации оксигемоглобина. С одной стороны, содержание СО2 влияет на внутриклеточный рН (эффект Бора). С другой, накопление СО2 вызывает образование карбаминовых соединений вследствие ее взаимодействия с аминогруппами гемоглобина. Эти карбаминовые соединения служат в качестве аллостерических эффекторов молекулы гемоглобина и непосредственно влияют на связывание О2. Низкий уровень карбаминовых соединений вызывает сдвиг кривой вправо и снижение сродства гемоглобина к О2, что сопровождается увеличение высвобождения О2 в тканях. По мере роста РаСО2 сопутствующее ему увеличение карбаминовых соединений сдвигает кривую влево, повышая связывание О2 гемоглобином. Органические фосфаты, в частности 2,3-дифосфоглицерат (2,3-ДФГ), образуются в эритроцитах в процессе гликолиза. Продукция 2,3- ДФГ увеличивается во время гипоксемии, что является важным механизмом адаптации. Ряд условий, вызывающих снижение О2 в периферических тканях, таких как анемия, острая кровопотеря, застойная сердечная недостаточность и т.д. характеризуются увеличением продукции органических фосфатов в эритроцитах. При этом уменьшается сродство гемоглобина к О2 и повышается его высвобождение в тканях. И наоборот, при некоторых патологических состояниях, таких как септический шок и гипофосфатемия, наблюдается низкий уровень 2,3-ДФГ, что приводит к сдвигу кривой диссоциации оксигемоглобина влево. Температура тела влияет на кривую диссоциации оксигемоглобина менее выражено и клинически значимо, чем описанные выше факторы. Гипертермия вызывает повышение Р50, т.е. сдвиг кривой вправо, что является благоприятной приспособительной реакцией не повышенный кислородный запрос клеток при лихорадочных состояниях. Гипотермия, напротив, снижает Р50, т.е. сдвигает кривую диссоциации влево. СО, связываясь с гемоглобином (образуя карбоксигемоглобин), ухудшает оксигенацию периферических тканей посредством двух механизмов. Во-первых, СО непосредственно уменьшает кислородную емкость крови. Во-вторых, снижая количество гемоглобина, доступного для связывания О2; СО снижает Р50 и сдвигает кривую диссоциации оксигемоглобина влево. Окисление части двухвалентного железа гемоглобина до трехвалентного приводит к образованию метгемоглобина. В норме у здоровых людей метгемоглобин составляет менее 3% общего гемоглобина. Низкий его уровень поддерживается внутриклеточными ферментными механизмами восстановления. Метгемоглобинемия может наблюдаться как следствие врожденной недостаточности этих восстановительных ферментов или образования аномальных молекул гемоглобина, резистентных к ферментативному восстановлению (например, гемоглобин М). Доставка кислорода (DО2) представляет собой скорость транспорта кислорода артериальной кровью, которая зависит от кровотока и содержания О2 в артериальной крови. Системная доставка кислорода (DО2), рассчитывается как: DO2 = СаО2 х Qt (мл/мин) или DO2 = ([(Hb) 1,34 % насыщения] + [0,0031PaO2 )Qt (мл/мин) = 20 мл О2/100 мл крови 5000 мл/мин = 1000 мл О2/мин (3.23). Доставку и потребление кислорода часто рассчитывают с учётом площади поверхности тела. При сердечном индексе, составляющем 3 л/(мин*м-2) (Qt делённый на площадь поверхности тела) нормальное значение DО2 = 540 мл/(мин м2). Если обычный показатель сердечного выброса составляет от 2,5 до 3,5 л/мин/м2, то нормальная величина DО2 колеблется от 520 до 720 мл/мин/м2. Существует тонкое сопряжение между артериальным содержанием О2, сердечным выбросом, тканевой утилизацией О2 и содержанием О2 в смешанной венозной крови. Некоторые заболевания, такие как РДСВ и сепсис, сопровождаются нарушением сопряжения между утилизацией О2 периферическими тканями и доставкой кислорода. Утилизация снижается, когда доставка падает ниже некоторого порога. Отношение между этими переменными выражается правилом Фика, которое устанавливает, что потребление О2 (объем в 1 мин) является произведение минутного сердечного выброса и артерио-венозной разницы О2: Потребление О2 = VO2 = Q (CaO2 - CvO2) (3.24). В условия основного обмена взрослый человек потребляет около 250 мл О2 в минуту, с учетом площади поверхности тела - 110-160 мл/(мин*м2). Однако скорость утилизации О2 различными тканями неодинакова. Содержание кислорода в смешанной венозной крови представляет собой усредненную величину для венозной крови от всех органов - и низким, и с высоким уровнями экстракции О2. Возросшая кислородная потребность при фиксированном минутном сердечном выбросе вызывает увеличение артерио-венозной разницы по О2. Кроме того, нормальный компенсаторный ответ на снижение кровотока проявляется также в виде увеличения поглощения кислорода, достаточного для поддержания VO2 на нормальном уровне. Иными словами, падение сердечного выброса компенсируется увеличением разницы SaO2 – SvO2, и VO2 остаётся неизменным. Следовательно, артериовенозную разницу можно рассматривать как меру адекватности доставки кислорода, а снижение SvO2 отражает увеличение экстракции кислорода. При нормальном потреблении кислорода около 250 мл/мин и сердечном выбросе 5000 мл/мин нормальная артериовенозная разница, согласно этому уравнению, составит 5 мл О2/100 мл крови. При этом нормальный коэффициент экстракции О2 [(СаО2 - CvO2)/CaO2] составит 25 %, т. е. 5 мл/20 мл. Таким образом, в норме организм потребляет только 25 % кислорода, переносимого гемоглобином. Когда потребность в О2 превосходит возможность его доставки, то коэффициент экстракции становится выше 25 %. Наоборот, если доставка О2 превышает потребность, то коэффициент экстракции падает ниже 25 %. Если доставка кислорода снижена умеренно, потребление кислорода не изменяется благодаря увеличению экстракции О2 (насыщение гемоглобина кислородом в смешанной венозной крови снижается). В этом случае VO2 не зависит от доставки. По мере дальнейшего снижения DO2 достигается критическая точка, в которой VO2 становится прямо пропорциональна DO2. Состояние, при котором потребление кислорода зависит от доставки, характеризуется прогрессирующим лактат-ацидозом, обусловленным клеточной гипоксией. Критический уровень DO2 наблюдается в различных клинических ситуациях. Например, его значение 300 мл/(мин*м2) отмечено после операций в условиях искусственного кровообращения и у больных с острой дыхательной недостаточностью. Напряжение углекислого газа в смешанной венозной крови (PvCO2) в норме составляет примерно 46 мм рт. ст., что является конечным результатом смешивания крови, притекающей из тканей с различными уровнями метаболической активности. Венозное напряжение углекислого газа в венозной крови меньше в тканях с низкой метаболической активностью (например, в коже) и больше в органах с высокой метаболической активностью (например, в сердце). Двуокись углерода легко диффундирует. Ее способность к диффузии в 20 раз превышает таковую у кислорода. СО2, по мере образования в процессе клеточного метаболизма, диффундирует в капилляры и транспортируется к легким в трех основных формах: в виде растворенной СО2, в виде аниона бикарбоната и в виде карбаминовых соединений. СО2 очень хорошо растворяется в плазме. Количество растворенной фракции определяется произведением парциального давления СО2 и коэффициента растворимости ( =0,3 мл/л крови /мм рт. ст.). Около 5% общей двуокиси углерода в артериальной крови находится в форме растворенного газа. Анион бикарбоната является преобладающей формой СО2 (около 90%) в артериальной крови. Бикарбонатный анион является продуктом реакции СО2 с водой с образованием Н2СО3 и ее диссоциации: СО2 + Н2О Н2СО3Н+ + НСО3- (3.25). Реакция между СО2 и Н2О протекает медленно в плазме и очень быстро в эритроцитах, где присутствует внутриклеточный фермент карбонгидраза. Она облегчает реакцию между СО2 и Н2О с образованием Н2СО3. Вторая фаза уравнения протекает быстро без катализатора. По мере накопления НСО3- внутри эритроцита анион диффундирует через клеточную мембрану в плазму. Мембрана эритроцита относительно непроницаема для Н+, как и вообще для катионов, поэтому ионы водорода остаются внутри клетки. Электрическая нейтральность клетки в процессе диффузии СО2 в плазму обеспечивает приток ионов хлора из плазмы в эритроцит, что формирует так называемый хлоридный сдвиг (сдвига Гамбургера). Часть Н+, остающихся в эритроцитах, забуферируется, соединясь с гемоглобином. В периферических тканях, где концентрация СО2 высока и значительные количества Н+ накапливаются эритроцитами, связывание Н+ облегчается деоксигенацией гемоглобина. Восстановленный гемоглобин лучше связывается с протонами, чем оксигенированный. Таким образом, деоксигенация артериальной крови в периферических тканях способствует связыванию Н+ посредством образования восстановленного гемоглобина. СО2 + Н2О + HbО2 → HbH+ + HCO3+ О2 (3.26). Это увеличение связывания СО2 с гемоглобином известно как эффект Холдейна. В легких процесс имеет противоположное направление. Оксигенация гемоглобина усиливает его кислотные свойства, и высвобождение ионов водорода смещает равновесие преимущественно в сторону образования СО2: О2 + НСО3- + HbН+ → СО2 + Н2О + HbО2 (3.27). |