Семинар 10. БИОЛОГИЯ. Семинар 10 Хромосомная теория наследственности. Полное и неполное сцепление генов. Генетика пола
 Скачать 111 Kb. Скачать 111 Kb.
|

|
Семинар 10 «Хромосомная теория наследственности. Полное и неполное сцепление генов. Генетика пола» 1. Механизмы генотипического определения пола у организмов разных биологических видов Пол организмов, размножающихся половым способом, определяется парой особых половых хромосом, тогда как остальные хромосомы называют аутосомами. Пол, особи которого имеют одинаковые половые хромосомы, называется гомогаметным. У особей гомогаметного пола при образовании гамет все они будут одинаковыми по половым хромосомам. Пол, у особей которого половые хромосомы разные, называется гетерогаметным. Соответственно и половые клетки этих особей будут двух типов, причем образовываться они будут в равном соотношении. У половых хромосом нет принципиальных отличий от аутосом: они, как и аутосомы, несут гены, которые контролируют развитие тех или иных признаков. Половые хромосомы принято обозначать буквами. У видов с гетерогаметным мужским полом одинаковые по размеру и форме половые хромосомы обозначают буквой X, а хромосому, имеющуюся только у самцов, - буквой Y. Такой тип наследования пола встречается у многих червей, моллюсков и членистоногих, а из позвоночных - у млекопитающих и человека. В хромосомном комплексе самок этих видов две X-хромосомы, а у самцов - X-хромосома и Y-хромосома. У видов с гетерогаметным женским полом хромосому, имеющуюся как у самок, так и у самцов обозначают буквой Z, а имеющуюся только у самок - буквой W. Самки этих видов имеют хромосомный комплекс с половыми хромосомами ZW, а самцы - с половыми хромосомами ZZ. Такой тип наследования пола имеет место, в частности, среди растений - у клубники, среди насекомых - у бабочек, а среди позвоночных - у рыб, пресмыкающихся и птиц. Медоносная плела относится к видам, у которых гомогаметным является женский пол, но пчелиная матка откладывает два типа яиц - оплодотворенные и неоплодотворенные. Из оплодотворенных яйцеклеток (генотип XX) развиваются самки (матка и рабочие пчелы), а из неоплодотворенных (одна X- хромосома, генотип X0) - самцы (трутни). В связи с этим организмы, образовавшиеся при обыкновенном половом размножении, - диплоидные, а самцы, развивающиеся в результате партеногенеза, - гаплоидные. В процессе индивидуального развития в клетках тела трутней восстанавливается диплоидный хромосомный набор, тогда как в половых клетках гаплоидное число хромосом сохраняется и при образовании сперматозоидов не уменьшается. Аналогичное явление встречается и у некоторых других насекомых. Х- хромосома намного длиннее, чем Y-хромосома, и содержит некоторые гены, которых нет в Y-хромосоме, в том числе и гены, кодирующие не связанные с проявлением пола белки. ♀XX, ♂X0 (пустынная саранча) ♀XX, ♂XY (дрозофила, млекопитающие) ♀XY, ♂XX (бабочки, птицы, рептилии) ♀X0, ♂XX (тля, моль) ♀2n, ♂n гапло-диплоидный тип (пчелы) На базе генетической информации в зависимости от второй хромосомы на 2-12 неделях эмбриогенеза развивается гонадный пол – яичники или семенники. Они выделяют соответственно эстрогены или андрогены, под действием которых формируется гормональный пол. Одновременно определяется морфо-физиологический пол (мужской или женский фенотип) 2. Наследование, сцепленное с полом. Примеры у человека Анализ наследования признака окраски глаз у дрозофилы в лаборатории Т. Моргана выявил некоторые особенности, заставившие выделить в качестве отдельного типа наследования признаков сцепленное с полом наследование. Зависимость результатов эксперимента от того, кто из родителей являлся носителем доминантного варианта признака, позволила высказать предположение, что ген, определяющий окраску глаз у дрозофилы, расположен в Х-хромосоме и не имеет гомолога в Y-хромосоме. Все особенности сцепленного с полом наследования объясняются неодинаковой дозой соответствующих генов у представителей разного— гомо- и гетерогаметного пола. Гомогаметный пол несет двойную дозу генов, расположенных в Х-хромосоме. Развитие соответствующих признаков у гетерозигот (ХAХa) зависит от характера взаимодействия между аллельными генами. Гетерогаметный пол имеет одну Х-хромосому (ХО или XY). У некоторых видов Y-хромосома генетически инертна, у других она содержит некоторое количество структурных генов, часть из которых гомологична генам Х-хромосомы. Гены негомологичных участков Х- и Y-хромосом (или единственной Х-хромосомы) у гетерогаметного пола находятся в гемизиготном состоянии. Они представлены единственной дозой: ХAY, ХaХ, XYB. Формирование таких признаков у гетерогаметного пола определяется тем, какой аллель данного гена присутствует в генотипе организма. Характер наследования сцепленных с полом признаков в ряду поколений зависит от того, в какой хромосоме находится соответствующий ген. В связи с этим различают Х-сцепленное и Y-сцепленное (голандрическое) наследование. Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х- хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству. У млекопитающих (в том числе и человека) мужской пол получает Х- сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям. Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери. Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы — только «дочерям». При Х-сцепленном наследовании, так же как и при аутосомном, возможен промежуточный характер проявления признака у гетерозигот. Например, у кошек пигментация шерсти контролируется Х-сцепленным геном, разные аллели которого определяют черную (XA и рыжую (XA’) пигментацию. Гетерозиготные самки XAXA’ имеют пеструю окраску шерсти. Самцы же могут быть либо черными (XAY, либо рыжими (XA’Y). Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну. У человека X-хромосома, которую мужчина получает от матери, несет гены дальтонизма (цветовой слепоты) и гемофилии (несвертываемости крови). Эти гены рецессивны; у женщин названные болезни проявляются крайне редко, а у мужчин чаще, так как в Y-xpoмосомах мужчин нет доминантного аллеля, подавляющего действие этих генов, Y-хромосома несет такие признаки, как, на пример, волосатость мочки ушей, перепонка между пальцами ног, поэтому эти признаки проявляются только у мужчин как носителей Y-хромосомы. Всего с полом сцеплено около 120 признаков. 3. Понятие о наследовании ограниченном полом и контролируемым полом Признаки, ограниченные полом: гены их наход-ся в аутосомах, т.е. есть у обоих полов, но проявляются только у одного пола. 1) Молочная продуктивность. 2) Яичная продуктивность. 3) Икра у рыб (у женского пола). 4) Яркое оперение (у самцов). К числу нежелательных, ограниченных полом признаков, относятся: 1) триторхизм, 2) аномалия спермиев (у самцов), 3) недоразвитие частей половых органов (у самок). Признаки, контролируемые полом: гены в аутосомах, т.е. есть у обоих полов и проявляются также у обоих, только у одного пола чаще или интенсивнее, чем у другого. 1) Комолость доминирует у овец, рецессивно у баранов. 2) Заращение яйцеводов и семяпроводов у птиц доминантно у самок и рецессивно у самца. 3) Атаксия (расстройство координации движения) доминантно у самок и рецессивно у самца. 4) Искривление киля у птицы доминантно у самцов и рецессивно у самок. 5) Наследственное облысение доминантно у мужчин и рецессивно у женщин. 6) Указательный палец длиннее безымянного доминантно у мужчин и рецессивно у женщин. 4. Полное и неполное сцепление генов. Группа сцепления В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование. Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде. 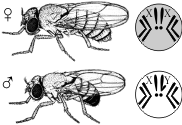 Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме. 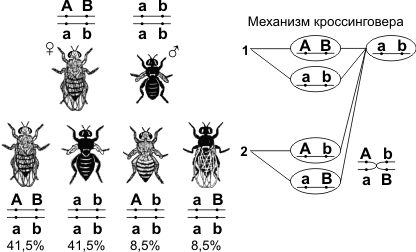 1 — некроссоверные гаметы; 2 — кроссоверные гаметы. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом. Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей. Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей. Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам. 5. Работы Т. Моргана по экспериментальному доказательству хромосомной теории Работы Моргана заложили основы хромосомной теории наследственности, они показали, что ограничения в свободной комбинаторике некоторых генов обусловлены расположением этих генов в одной хромосоме и их физическим сцеплением. Морганом было установлено, что сцепление генов, расположенных в одной хромосоме, не является абсолютным. Во время мейоза хромосомы одной пары могут обмениваться гомологичными участками между собой с помощью процесса, который называется кроссинговером. Чем дальше друг от друга расположены гены в хромосоме, тем чаще они разделяются кроссинговером. На основе этого феномена была предложена мера силы сцепления генов — процент кроссинговера — и построены первые генетические карты хромосом для разных видов дрозофилы. В качестве объекта генетического анализа была выбрана плодовая мушка дрозофила и Морган изучал наследование у нее разных признаков. Скрестив гомозиготную самку с серыми телом и длинными крыльями (домин), с гомозиготным чернокрылым короткокрылым самцом, в F1 – однообразие (серое тело, длинные крылья) Далее Морган провел несколько анализирующих скрещиваний. Оказалось, что результаты будут разные в зависимости от пола гибрида. Если гибридным был самец, то в потомстве получалось 2 фенотипических класса полностью повторяющих признаки родителей. Если гибридной была самка, то получалось 4 фенотипических классов потомком в неравных пропорциях. Большую часть потомства (83%) составляют потомки с родительскими признаками, меньшую (17%) – особи с новыми комбинациями признаков. Морган сделал вывод, что сцепление может быть неполным, где группа сцепления нарушается кроссинговером. Необычность процентного соотношения у потомков объясняется тем, что кроссинговер происходит не всегда, частота кроссинговера зависит от расстояния между генами – чем больше расстояние, тем меньше силы сцепления между генами, тем чаще кроссинговер. Гаметы, в которые попали хромосомы, не прошедшие кроссинговер, называются некроссоверные. Если в гаметах хромосомы претерпевшие кроссинговер – кроссоверные. 6. Основные положения хромосомной теории наследственности 1. Гены расположены в хромосомах линейно в определенных участках – локусах. Аллельные гены занимают одинаковые локусы гомологичных хромосом. 2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе или сцеплено. Число групп сцепления = числу хромосом в гаплоидном наборе. 3. Между гомологичными хромосомами возможен кроссинговер, нарушающий сцепление 4. процесс кроссинговера прямо пропорционален расстоянию между генами. 1% кроссинговера = 1 сантиморганида 7. Понятие о цитоплазматической наследственности Наличие некоторого количества наследственного материала в цитоплазме в виде кольцевых молекул ДНК митохондрий и пластид, а также других внеядерных генетических элементов дает основание специально остановиться на их участии в формировании фенотипа в процессе индивидуального развития. Цитоплазматические гены не подчиняются менделевским закономерностям наследования, которые определяются поведением хромосом при митозе, мейозе и оплодотворении. В связи с тем что организм, образуемый вследствие оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был впервые описан в 1908 г. К. Корренсом в отношении признака пестрых листьев у некоторых растений. |
