Физиология растений и животных. Скопичев В. Г. Физиология растений и животных Направление подготовки 020400 биология Профиль подготовки Биоэкология
 Скачать 35.41 Mb. Скачать 35.41 Mb.
|

10.1.6. Аккомодация.При медленном нарастании силы тока возбуждение не наступает. Это явление получило название аккомодации и связано с тем, что повышение мембранного потенциала к пороговому уровню вызывает открывание натриевых каналов и соответственно возникновение входящего натриевого тока. Однако если этот процесс происходит с недостаточной скоростью, часть натриевых каналов успевает закрыться. В то же время происходит активация калиевых каналов, у которых, в отличие от натриевых, инактивации со временем не происходит. Таким образом, ослабляется входящий натриевый ток и усиливается выходящий калиевый ток, что замедляет или блокирует развитие регенеративного процесса (закон градиента). 10.1.7. Закон длительности раздражения.Показано, что порог возбуждения не достигается, если раздражение длится очень долго, но сила его мала, или сила раздражения велика, но длится очень короткий промежуток времени. Начиная с пороговой силы, время становится значимым (полезным) для раздражения. По существу «полезное время» - это критическая длительность действия наименьшего тока, способного вызвать возбуждение. Для того чтобы импульс тока мог изменить мембранный потенциал и вызвать возникновение потенциала действия, через неё должно пройти некоторое минимальное количество электричества (Q), которое как известно измеряется как произведение силы тока (I) на время (t) Q = It. Из соотношения следует, что по мере сокращения длительности тока необходимо увеличить его силу, чтобы он сохранял эффективность в качестве раздражителя. Для импульсов большой длительности существует некоторая минимальная сила тока, достаточная для возбуждения. Более слабый ток из-за того, что мембрана в покое имеет определенную проводимость для ионов калия, не сможет вызвать необходимое смещение трансмембранной разности потенциалов. Чем короче полезное время, тем более возбудимыми являются ткани и быстрее реагируют на раздражитель. Точное измерение полезного времени затруднительно, поскольку в этой части кривой значительные изменения времени соответствуют очень малым изменениям порога раздражения. 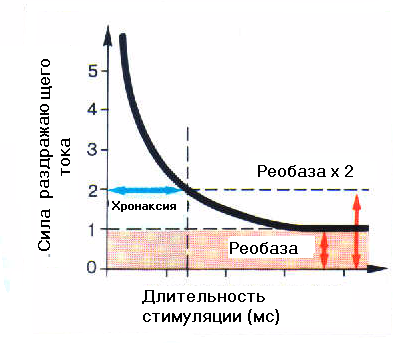 Принято измерять в качестве порога времени, минимальную длительность раздражителя при его силе, равным двум порогам и при условии, что сила порогового раздражителя измерена при длительностях времени большим, чем полезное время. Пороговая сила была обозначена как реобаза, а необходимое время раздражения при силе раздражения равным двум реобазам – хронаксия. Хронаксия – величина переменная и зависит от многих факторов: от структуры ткани, её функционального состояния и всего организма в целом. Кривые «сила – длительность» для разных тканей и органов подобны по форме, но в разных масштабах. Например, у лошади и жвачных животных хронаксия двигательных нервов колеблется от 0,09 до 0,2 мс, а скелетных мышц от 0,2 до 0,4 мс. Самая большая хронаксия, измеряемая не мс, а десятками и сотнями мс у гладких мышц желудка, кишечника и матки. 10.1.8. Распространение возбуждения.Нервные клетки способны передавать возникшее в них возбуждение другим клеткам. Несмотря на большое разнообразие нервных клеток в организме, сообщаются они между собой с помощью лишь двух основных типов электрических сигналов: 1) медленно изменяющихся, 2) импульсных потенциалов с определённой амплитудой, возникающих по закону «все или ничего» - потенциалов действия. Распространение возбуждения в нервных клетках происходит по их отросткам – нервным волокнам (аксонам или дендритам). Немиелинизированные нервные волокна покрыты мембраной, которая проводит электрический ток значительно хуже – аналог изолирующей оболочки кабеля. Идеальным изолятором является миелиновая оболочка мякотных волокон, но она прерывается участками проводящей клеточной мембраны через 0,9 – 2 мм. При подаче электрического сигнала на нервное волокно электрический ток распространяется по нему в соответствии с его электрическим сопротивлением ионному току цитоплазмы и мембраны. Для миелинизированного нервного волокна сопротивление волокна на участке, покрытом миелином, имеет очень большую величину по сравнению с сопротивлением мембраны перехвата Ранвье. 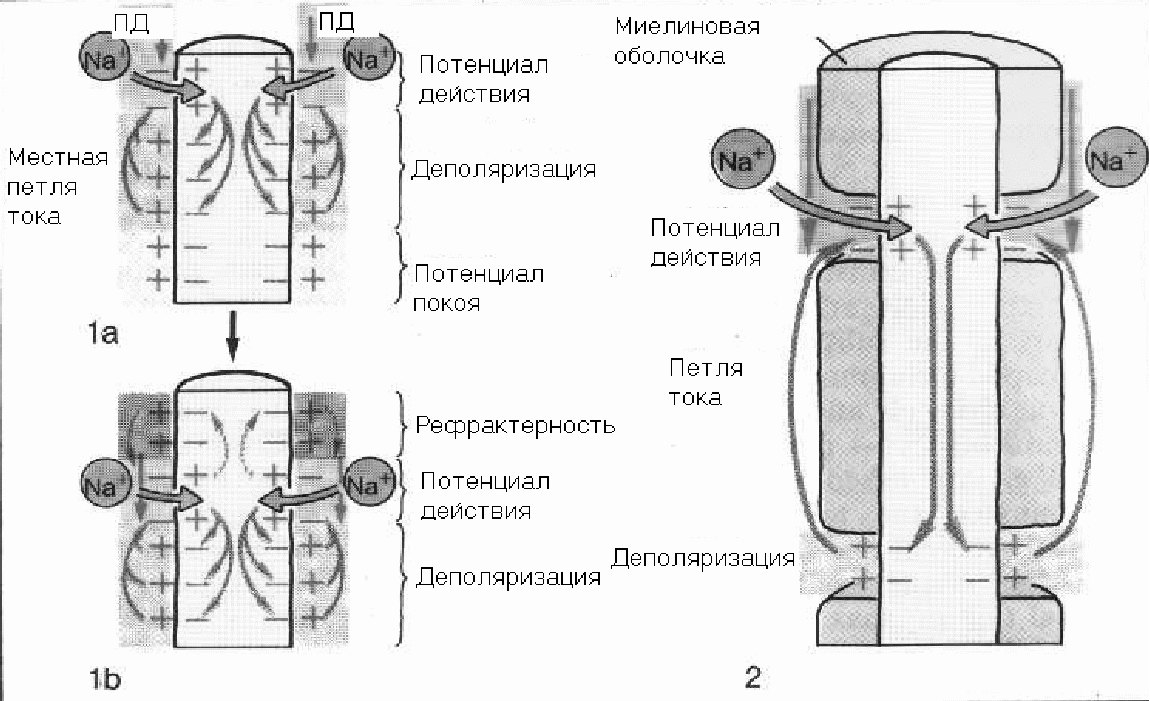 Электрический ток будет распространяться с меньшими потерями вдоль миелинизированного нервного волокна, поскольку изолирующие свойства миелиновой оболочки снижают утечку тока через поверхность волокна. В этих случаях клетки используют для распространения возбуждения регенеративный механизм генерации электрического возбуждения. Электрический ток будет распространяться с меньшими потерями вдоль миелинизированного нервного волокна, поскольку изолирующие свойства миелиновой оболочки снижают утечку тока через поверхность волокна. В этих случаях клетки используют для распространения возбуждения регенеративный механизм генерации электрического возбуждения. В результате воздействия какого-либо внешнего или внутреннего источника раздражения определённый участок нервного волокна возбуждается и лавинообразно увеличивается проводимость для ионов натрия, а внутренняя поверхность мембраны становится на время положительной. Ионный ток будет распространяться от положительного полюса к отрицательному, а петли тока будут проходить вдоль нервного волокна по цитоплазме и через мембрану невозбуждённых участков. В результате мембранный потенциал будет сдвигаться к пороговому значению и в случае его превышения проницаемость ионов натрия начнет регенеративно увеличиваться с последующим возникновением потенциала действия. Появление потенциала действия в новом участке вызывает местные токи деполяризующие и возбуждающие следующие участки нервного волокна. В миелинизированном волокне он будет несколько отличаться. В частности, из-за того, что участки мембраны между перехватами Ранвье покрыты миелином, весь возбуждающий местный ток будет воздействовать только на мембрану в области перехвата Ранвье, и возникновение очередного потенциала действия будет происходить только в последующем перехвате. Если распространение в безмякотном нервном волокне происходит плавно и напоминает движение горящего участка в бикфордовом шнуре, то в мякотном волокне возбуждение будет распространяться скачкообразно или как часто используют термин – сальтаторно от перехвата к перехвату. Таким образом, распространяющийся по нервному волокну сигнал постоянно усиливается и поддерживается на одинаковом уровне. Порог деполяризации для возбуждения мембраны составляет около 20 мВ, тогда как обычная деполяризация мембраны при максимальной амплитуде потенциала действия равна 100 – 120 мВ. Следовательно, при генерации потенциала действия происходит усиление сигнала в 5-6 раз. Скорость проведения возбуждения зависит от диаметра волокна, а также от наличия у нервного волокна миелиновой оболочки. У высших позвоночных животных наименьшая скорость проведения - 0,5- 2 м/с. была зарегистрирована в тонких немиелинизированных волокнах диаметром 0,5-1,0 мкм. Как правило, это вегетативные нервные волокна. Наибольшей скоростью (70 – 120 м/с) обладают самые толстые миелинизированные волокна диаметром 15 – 25 мкм. К ним относятся, например, чувствительные волокна особенно передающие болевые раздражения и Иннервирующие ряд скелетных мышц. Миелинизированные нервные волокна при равных диаметрах с немиелинизированными имеют большую скорость проведения. Плотность натриевых каналов в мембране перехвата Ранвье в несколько раз выше, чем в мембране немиелинизированных волокон. Это дополнительно способствует более быстрой деполяризации мембраны в области перехвата Ранвье и соответственно увеличивает скорость проведения по волокну. |