Физиология растений и животных. Скопичев В. Г. Физиология растений и животных Направление подготовки 020400 биология Профиль подготовки Биоэкология
 Скачать 35.41 Mb. Скачать 35.41 Mb.
|

10.1.8.1. Рефрактерность.После окончания возбуждения в нервных или мышечных клетках или, другими словами, после окончания в них потенциала действия наступает временное состояние невозбудимости – рефрактерности. После сокращения сердца очередное сокращение нельзя было вызвать в течении периода, равного десятым долям секунды независимо от амплитуды и длительности раздражающего стимула. В нервных клетках период невозбудимости оказался значительно короче. При уменьшении интервала раздражения между двумя раздражающими электрическими стимулами величина потенциала действия в ответ на второй стимул становится все меньше и меньше. А если повторный стимул наносится во время генерации потенциала действия или сразу же после его окончания, второй потенциал действия не генерируется. Период, в течении которого, потенциал действия на второй раздражающий стимул не возникает, получил название абсолютного рефрактерного периода. Он составляет для нервных клеток позвоночных животных 1,5 – 2 мс. После периода абсолютной рефрактерности наступает относительный рефрактерный период. Он характеризуется: 1) повышенным порогом раздражения по сравнению с исходным состоянием (т.е. для того чтобы возник повторный потенциал действия, необходим ток большей величины) 2) снижением амплитуды потенциала действия. По мере окончания периода относительной рефрактерности возбудимость повышается до исходного уровня, и величина порогового раздражения уменьшается также до первоначального значения. В период абсолютной рефрактерности наблюдается повышенная калиевая проводимость за счёт открывания дополнительных калиевых каналов и снижение натриевой проводимости за счёт инактивации натриевых каналов. Поэтому даже при больших значениях деполяризующего тока не удаётся активировать такое количество натриевых каналов, чтобы выходящий натриевый ток мог бы превысить увеличенный выходящий калиевый ток и снова запустить регенеративный процесс. Во время относительного рефрактерного периода деполяризующий сигнал достаточно большой амплитуды может активировать воротный механизм натриевых каналов так, что несмотря на большое число открытых калиевых каналов натриевая проводимость увеличивается и вновь возникает потенциал действия. Вместе с тем из-за увеличенной проводимости мембраны к ионам калия и остаточной натриевой инактивации повышение мембранного потенциала не будет уже столь близко к значению равновесного натриевого потенциала. Поэтому потенциал действия будет меньшим по амплитуде. Далее следует фаза экзальтации – повышенной возбудимости возникающей в результате, наличия следовой деполяризации. В последующем при развитии следовой гиперполяризации наступает фаза субнормальности – характеризующаяся снижением амплитуды потенциалов действия. Наличие рефрактерных фаз обуславливает прерывистый (дискретный) характер нервной сигнализации, а ионный механизм генерации потенциала действия обеспечивает стандартность нервных импульсов. Вследствие этого изменения внешних сигналов кодируются изменением частоты потенциалов действия. Максимально возможный ритм активности, лимитированный длительностью абсолютной рефрактерной фазы обозначают как лабильность (функциональную подвижность). У нервных волокон лабильность составляет 200 - 400 Гц, а у некоторых чувствительных нервных волокон достигает 1кГц. В случае, когда новый раздражающий импульс приходится на фазу экзальтации реакция ткани становится максимальной – развивается оптимум частоты. При попадании последующего стимулирующего импульса на фазу относительной или абсолютной рефрактерности реакция ткани ослабляется или прекращается вовсе, развивается пессимальное торможение. 10.1.8.2 Передача нервного возбуждения между клетками. Представление о синапсах.Если клеточная мембрана нервного отростка примыкает к мембране другой нервной клетки или к мембране мышечного мышечного волокна то от этого участка через мембрану отростка и мембрану другой клетки будут пассивно распространяться петли тока деполяризуя мембрану соседней клетки. При достижении порогового значения в ней начнётся процесс повышения проводимости к ионам натрия и возникнет потенциал действия. Такое соединение получило название электрического синапса. Для того чтобы передача функционировала более эффективно необходимо увеличить площадь соприкосновения двух мембран, а также плотнее приблизить сами мембраны. Электрическая передача возбуждения имеет определённые преимущества, поскольку ионный ток при такой передаче непосредственно течёт из передающей (пресинаптической) клетки в воспринимающую (постсинаптическую) клетку без каких-либо промежуточных этапов, задержка при проведении возбуждения будет минимальная, и нужно быстро охватить возбуждением нескольких нервных клеток. В 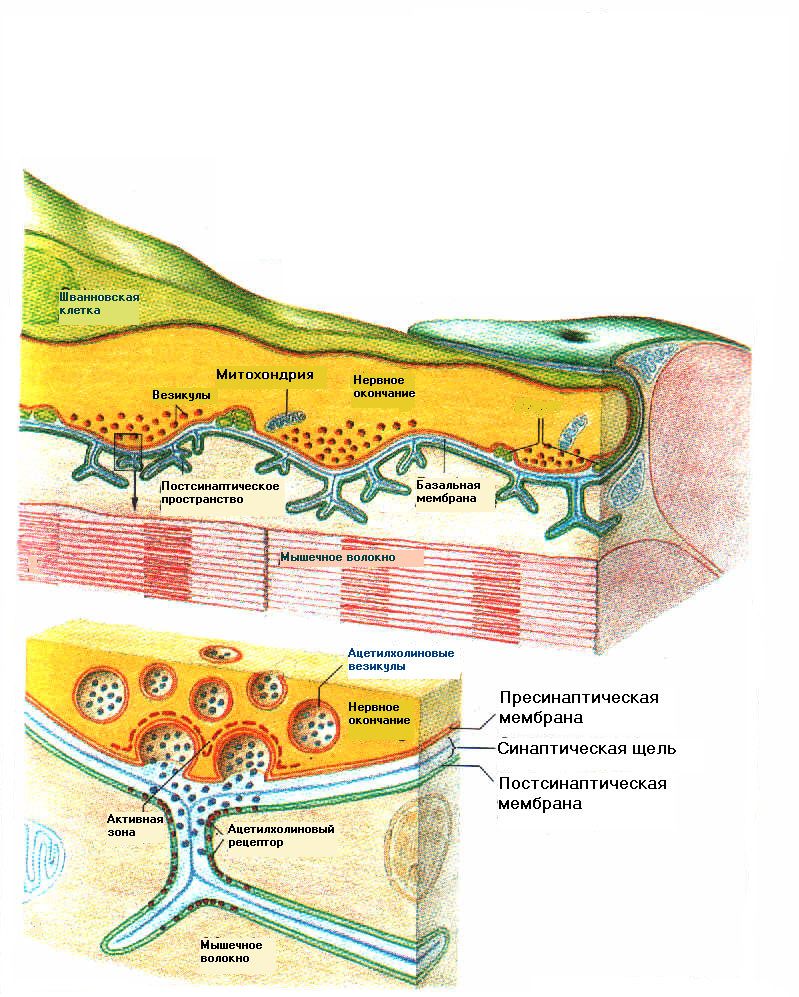 большинстве синапсов используются специальные химические вещества посредники – медиаторы. После возникновения потенциала действия в пресинаптическом окончании мембрана его деполяризована. При этом помимо натриевых и калиевых каналов активируются (открываются) кальциевые каналы и в окончание из внешней среды входят ионы кальция. Повышение внутриклеточной концентрации кальция вызывает выброс медиатора из везикул. Медиатор поступает во внеклеточное пространство и диффундирует к постсинаптической мембране связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация хемичувствительных ионных каналов и возникает постсинаптический ток, под действием которого развивается потенциал действия мышечной клетки. Так в нервно-мышечном синапсе функционирует ацетилхолин (АцХ) - активируемый канал– интегральный белок мышечной мембраны, состоит из пяти белковых субъединиц, которые образуют трубчатую структуру с общей молекулярной массой около 250000. Рецепторный участок канала состоит из двух белковых субъединиц и располагается на наружной поверхности мембраны. АцХ каналы и связанные с ними АцХ рецепторы сосредоточены в постсинаптической мембране концевой пластинки и плотность их весьма велика – около 104/мкм2. большинстве синапсов используются специальные химические вещества посредники – медиаторы. После возникновения потенциала действия в пресинаптическом окончании мембрана его деполяризована. При этом помимо натриевых и калиевых каналов активируются (открываются) кальциевые каналы и в окончание из внешней среды входят ионы кальция. Повышение внутриклеточной концентрации кальция вызывает выброс медиатора из везикул. Медиатор поступает во внеклеточное пространство и диффундирует к постсинаптической мембране связывается со специальными (рецепторными) участками этой мембраны. В результате происходит активация хемичувствительных ионных каналов и возникает постсинаптический ток, под действием которого развивается потенциал действия мышечной клетки. Так в нервно-мышечном синапсе функционирует ацетилхолин (АцХ) - активируемый канал– интегральный белок мышечной мембраны, состоит из пяти белковых субъединиц, которые образуют трубчатую структуру с общей молекулярной массой около 250000. Рецепторный участок канала состоит из двух белковых субъединиц и располагается на наружной поверхности мембраны. АцХ каналы и связанные с ними АцХ рецепторы сосредоточены в постсинаптической мембране концевой пластинки и плотность их весьма велика – около 104/мкм2. Проведение в химическом синапсе, из-за наличия химического звена, происходит медленнее, чем в электрическом. Однако химическая передача более гибкая, чем электрическая, поскольку с её помощью легко может осуществляться как возбуждающее, так и тормозное действие. Кроме того, при активации постсинаптических каналов химическими агентами может возникать достаточно сильный ток, способный деполяризовать до порогового уровня крупные клетки. В связи с этим при химической передаче тонкие пресинаптические волокна могут возбуждать большие постсинаптические клетки. После окончания действия медиатора он разрушается специальными ферментами. В случае, холинергических синапсов медиатор ацетилхолин гидролизуется холинэстеразой. |