Занятие №9МП. Сложные липиды. Пол
 Скачать 1.66 Mb. Скачать 1.66 Mb.
|

|
Сложные липиды. ПОЛ(Пероксидное окисление липидов в клеточных мембранах) ЧАСТЬ 1. Сложные омыляемые липиды (в отличие от простых) содержат кроме спирта и жирной кислоты дополнительные компоненты (см. схему). Комплексы липидов и белков называются ПРОТЕОЛИПИДАМИ и встречаются в липопротеинах и мембранах. 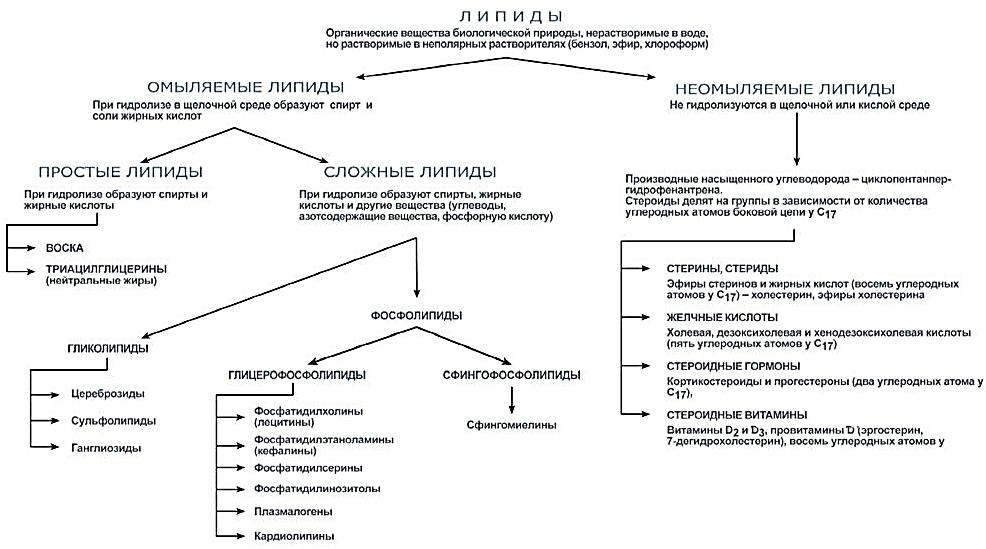 В роли спирта глицерин присутствует в глицерофосфолипидах, сфингозин (аминоспирт) - в гликолипидах и сфингофосфолипидах (иногда их объединяют и называют сфинголипидами). 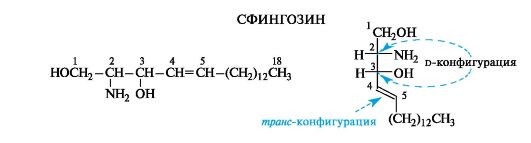 Простыми сфинголипидами являются церамиды. Они содержат только сфингозин, соединённый с жирнокислотным ацильным остатком. Продукт взаимодействия сфингозина и жирной кислоты называется церамид (N-ацилсфингозин). 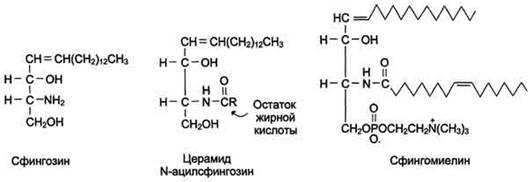 1.1.Фосфолипиды – сложные липиды, содержащие, помимо жирных кислот и спирта (глицерина или сфингозина), остаток фосфорной кислоты. Фосфолипидыесть во всех живых клетках. Содержатся в нервной ткани, участвуют в доставке жиров, жирных кислот и холестерина. Наряду с холестерином фосфолипиды являются структурными компонентами биомембран, а благодаря дифильности фосфолипиды образуют поверхностный гидрофильный слой липопротеинов крови, выстилают поверхность альвеол, предотвращая слипание стенок во время выдоха. Биологическая роль фосфолипидов: «Растворители» холестерина. Приостанавливают развитие атеросклероза. Обладают антиоксидантным действием. Ценное противоопухолевое средство. Защищают печень при использовании анаболических стероидов и антибиотиков (гепатопротеторное действие). Дефицит фосфолипидов приводит: к развитию жировой инфильтрации печени; к нарушению желчеобразования и желчевыделения; к увеличению проницаемости клеточных мембран. 1.1.1. В состав глицеролфосфолипидов входят фосфатидная кислота (ее кислотный остаток называется фосфотидил) и обычно азотсодержащие соединения (этаноламин, холин, серин) или циклический шестиатомный спирт циклогексана – инозит (инозитол). Фосфатидная кислота - продукт последовательных реакций этерификации глицерина, фосфорной кислоты и двух жирных кислот. 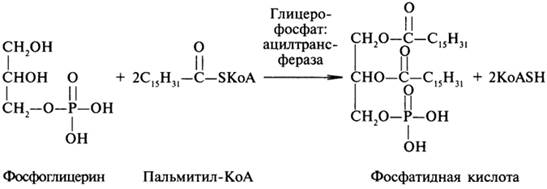 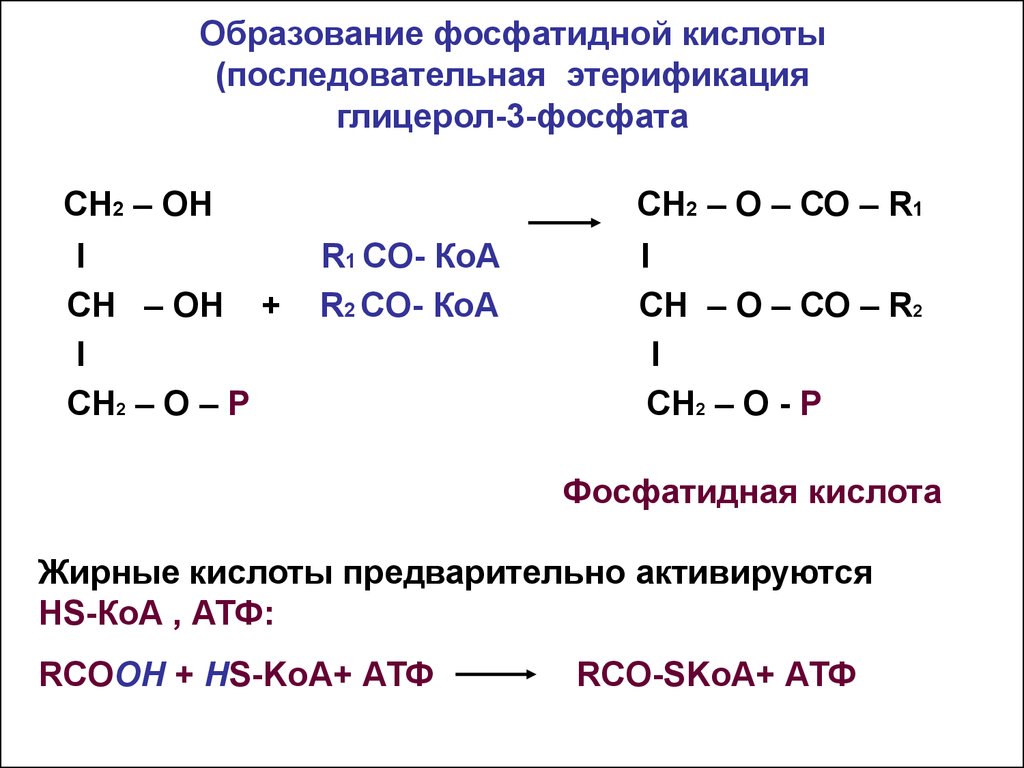 Фосфатидная кислота в свободном состоянии в организме содержится в большом количестве. Фосфатидная кислота может содержать насыщенные или ненасыщенные жирные кислоты. Жирнокислотный состав фосфатидной кислоты определяет ее дальнейшую судьбу: - если используются насыщенные и мононенасыщенные кислоты (пальмитиновая, стеариновая, пальмитолеиновая, олеиновая), то фосфатидная кислота направляется на синтез триацилглицеролов, - при включении полиненасыщенных жирных кислот (линоленовая, арахидоновая, кислоты ω3-ряда) фосфатидная кислота является предшественником глицерофосфолипидов. 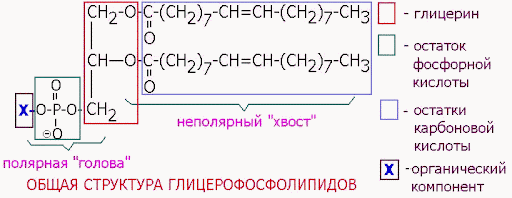 – 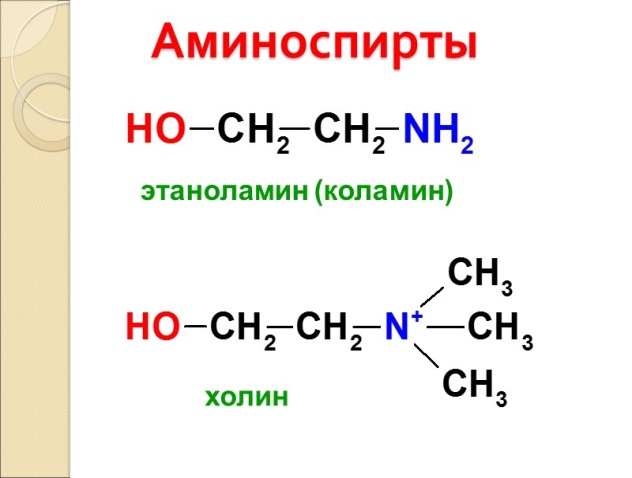 - холин,  - серин,  - инозитол Этаноламин всегда присутствует в организме в условиях нормального белкового питания, образуется при декарбоксилировании серина. Дальнейшие пути превращений связаны либо с синтезом одного из фосфатидов — кефалина (фосфатидилэтаноламина), либо с превращением этаноламина в холин. Инозитол называли "витамином В8", однако было показано, что около 3/4 суточной потребности инозитола вырабатывается самим организмом, поэтому инозитол относят к витаминоподобным веществам. Основные глицеролфосфолипиды в организме человека 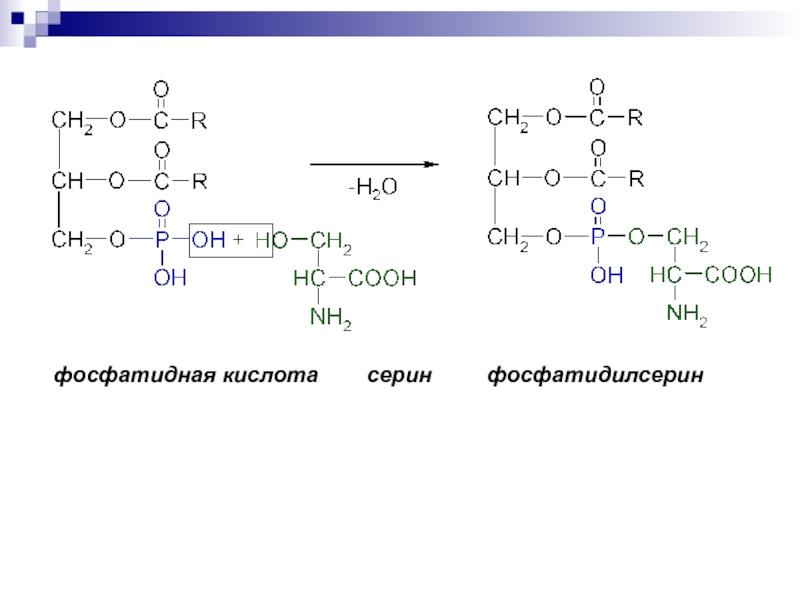 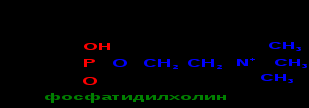 Аналогичным образом образуются: фосфотидилхолит (лецитин) Основными липидами мембран животных клеток являются: фосфатидилхолин и фосфатидилэтаноламин. Дипальмитинфосфотидилхолин – ПАВ, основной компонент сурфактанта легочный альвеол. Предотвращает слипание стенок во время выдоха. 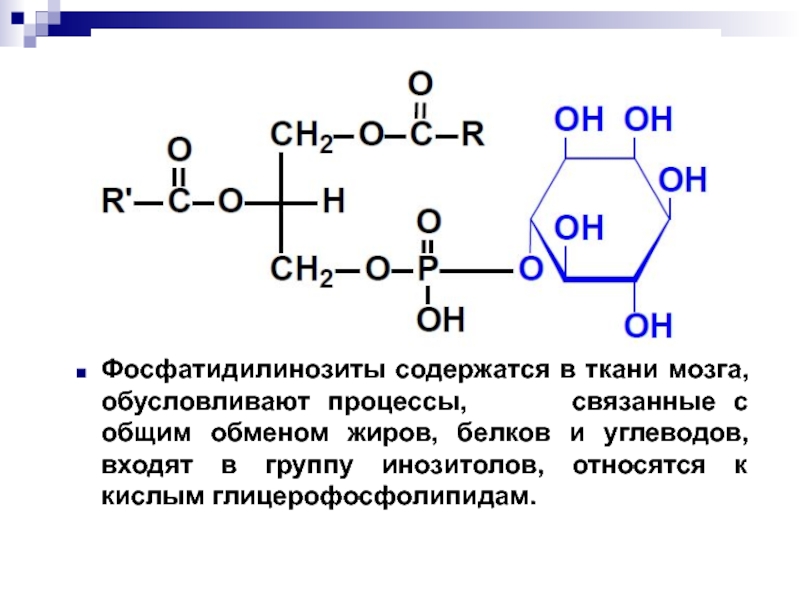 фосфатидилинозит Содержится в ткани мозга, обуславливает процессы, связанные с общим обменом жиров, белков и углеводов. Играет важную роль в фосфолипид-кальциевом механизме передачи гормонального сигнала в клетку. 1.1.2.Сфингомиелины содержат заряженную полярную группу, такую как фосфохолин или фосфоэтаноламин. 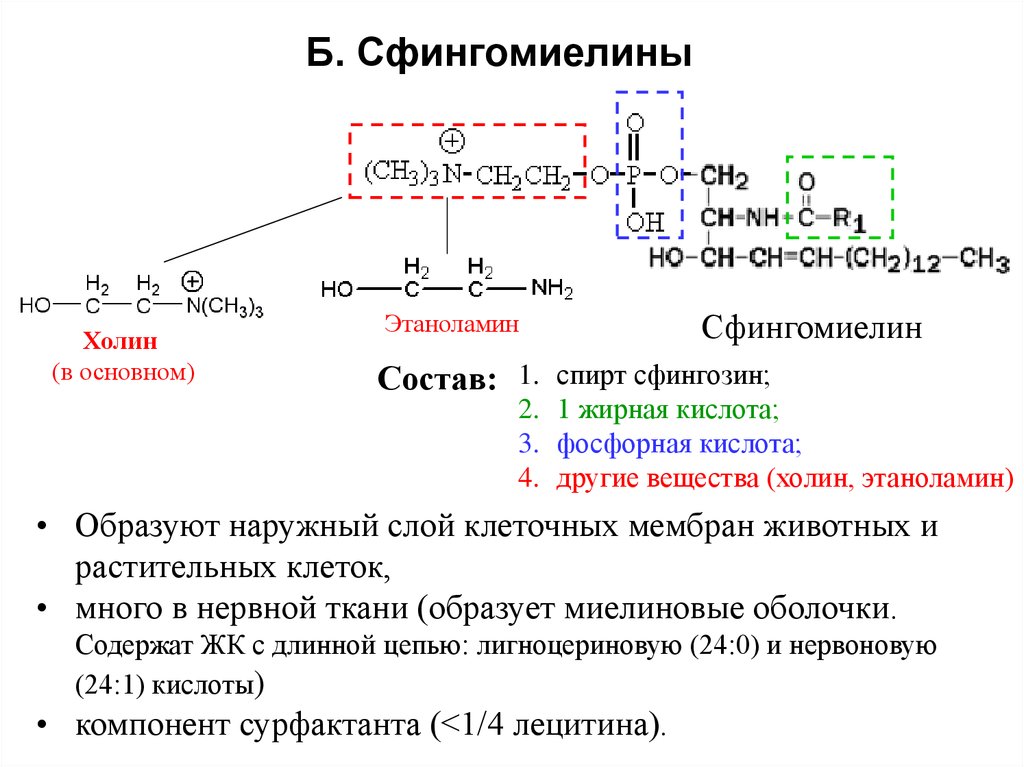 Дифильность сфингомиелина 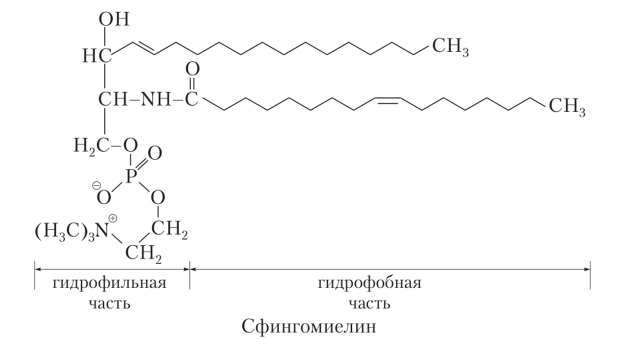 Сфингомиелины образуют наружный слой клеточных мембран животных и растительных клеток. Много в нервной ткани (образует миелиновые оболочки). В сером веществе мозга до 70% жирных кислот сфингомиелина представлено стеариновой кислотой. 1.2.Гликолипиды (гликосфинголипиды) содержат церамид, этерифицированный по 1-гидрокси-группе остатком сахара. В зависимости от сахара гликосфинголипиды подразделяются на сульфатиды, цереброзиды и ганглиозиды. Заметим, что в них отсутствует фосфатная группа. 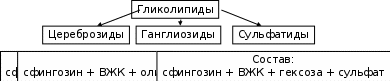 1.2.1. Цереброзиды содержат в качестве остатка сахара глюкозу или галактозу (глюкозилцерамид и галактозилцерамид). Находятся преимущественно в белом веществе головного мозга. 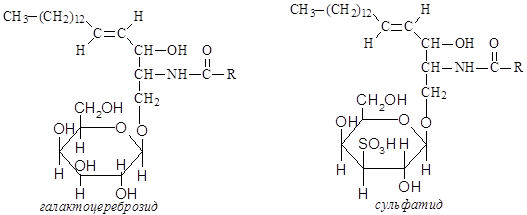 1.2.2 Сульфатиды - соединения с сульфогруппой на углеводных остатках. 1.2.3. Ганглиозиды — представители наиболее сложно построенных гликолипидов. Содержат трисахарид, причём один из них всегда сиаловая кислота. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа – углеводный остаток. 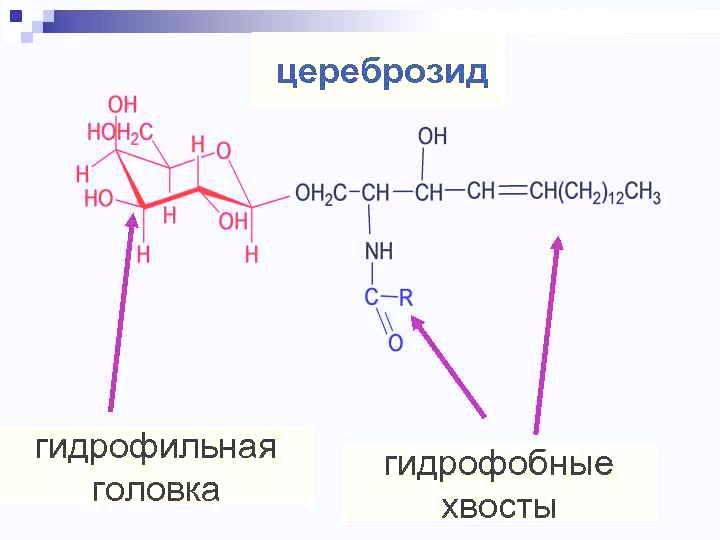 глюкоза остаток церамида Часть 2. Строение мембраны и липопротеинов крови.Протеолипиды (липопротеины) представляют собой комплексы липидов и белков, в которых доля липидов является преобладающей, белок включает много гидрофобных аминокислот. Связь между липидами и белками осуществляется посредством гидрофобных и электростатических взаимодействий. Наиболее изучены липопротеины плазмы крови, которые классифицируют по их плотности. Чем выше содержание в них липидов, тем ниже плотность липопротеинов. Различают липопротеины высокой плотности (ЛВП), низкой плотности (ЛНП), очень низкой плотности (ЛОНП) и хиломикроны. Все группы липопротеинов плазмы содержат полярные и неполярные липиды в разных соотношениях. Липопротеины осуществляют транспорт липидов в организме биологических жидкостей. Причем каждый из четырех классов липопротеинов выполняет свою роль в транспорте липидов. Строение мицеллы липопротеинов.  Липопротеины, это сферические частицы, в которых можно выделить гидрофобную серцевину, состоящую из триглицеридов и эфиров холестерина, и амфифильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки. Белки оболочки (апобелки) – стабилизируют жировые комплексы и позволяют им растворяться в плазме крови. Хиломикроны — класс липопротеинов, образующихся в тонком кишечнике в процессе всасывания экзогенных липидов. Это самые большие из липопротеинов, Образующиеся хиломикроны состоят на 85 % из триглицеридов, поэтому вместе с липопротеинами очень низкой плотности их относят к триглицерид-богатым липопротеинам. Кроме триглицеридов хиломикроны содержат также холестерин и эфиры холестерина. Протеин частично покрывает поверхность хиломикрона и, таким образом, обеспечивает стабильность частицы в процессе циркуляции. МЕМБРАНЫ КЛЕТОК 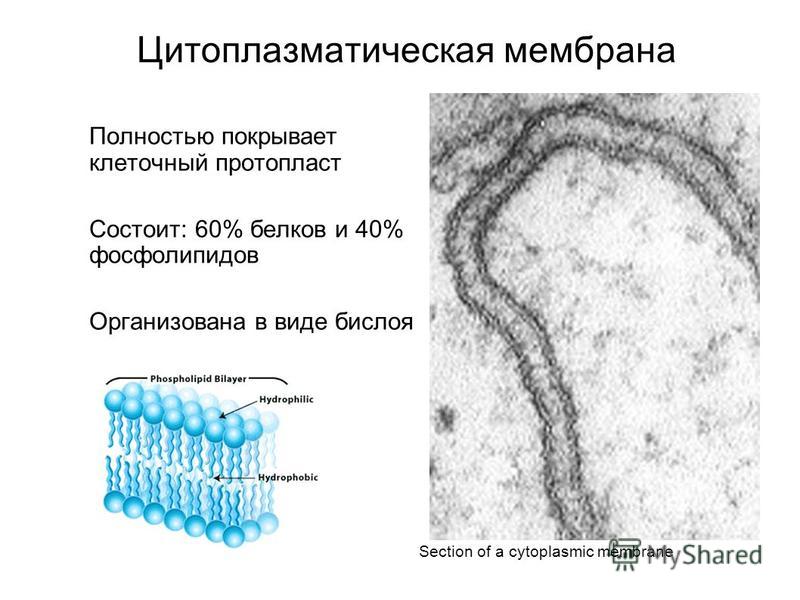 К клеточным мембранам относятся цитоплазматическая мембрана, ядерная мембрана, мембраны митохондрий, эндоплазматической сети и др. Все клеточные мембраны построены из молекул липидов и белков, толщиной не более 10 нм. В целом, все биомембраны имеют близкую структуру. При этом различаются по химическому составу липидов в бислое и по типу встроенных белков. Хотя соотношение белков и липидов в различных мембранах варьирует, в среднем, плазматические мембраны содержат 40% липидов, 50% белков и 5-10% углеводов. Липидный бислой – основной компонент, формирующий структуру мембраны, который представлен полярными липидами (фосфолипидами), стеринами и гликолипидами. Мембранные фосфолипиды дифильны и содержат гидрофобные части (алифатические радикалы жирных кислот), которые направлены вовнутрь мембран, и гидрофильные (полярные «головки»), располагающиеся на поверхности мембраны. Расположение белков в липидном бислое. Белки в липидных слоях мембран своими неполярными (гидрофобными) участками погружены в «жирную» часть мембраны, где находятся гидрофобные участки липидов. Гидрофобная часть белковой молекулы погружения в липидную часть мембраны, а аолярная (гидрофильная) часть таких белков взаимодействует с головками липидов и обращена напружу, в сторону водной фазы. Поэтому мембранные белки, связанные с липидами путем гидрофобных взаимодействий, прочно связываются с мембранами и практически не экстрагируются в водных фазах. 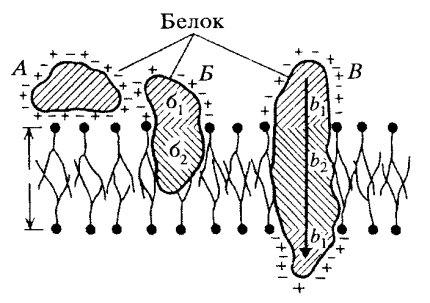 А-ионные связи, b1 - гидрофильное взаимодействие , b2 -гидрофобное взаимодействие А-ионные связи, b1 - гидрофильное взаимодействие , b2 -гидрофобное взаимодействиеСВОЙСТВА КЛЕТОЧНЫХ МЕМБРАН 1. Полупроницаемость 2. Механическая прочность 3. Текучесть. 4. Пластичность. 5. Ассиметричность 6. Самоорганизация. 7. Замкнутость. 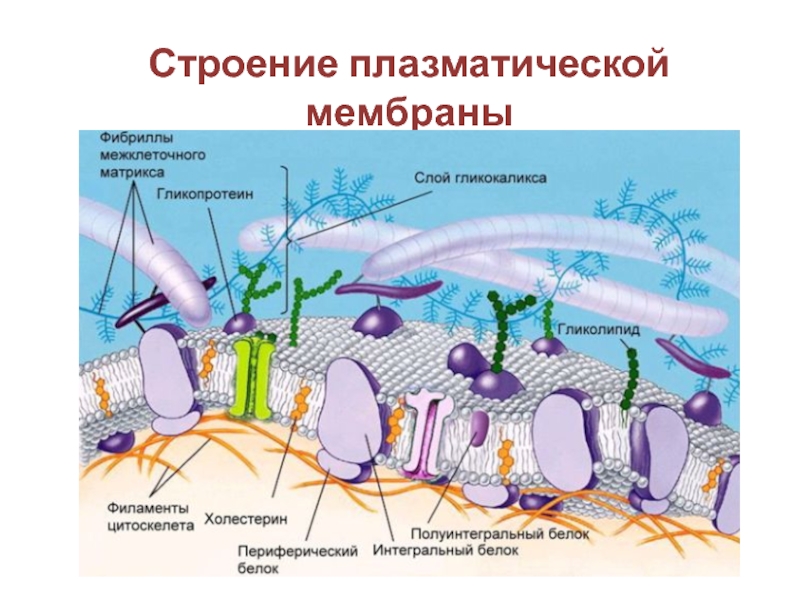 Часть 3. Пероксидное окисление липидов (ПОЛ) - это биофизический процесс обновления и перестройки биологических мембран, регуляции их состава, проницаемости и активности мембраносвязанных ферментов. Наиболее чувствительными компонентами клетки являются фосфолипиды мембран: в первую очередь, ненасыщенные жирные кислоты. Действующими агентами ПОЛ являются свободные радикалы, которые имеют в своей молекуле неспаренные электроны. Инициаторами ПОЛ в живом организме являются активные формы кислорода (АФК). Они возникают при недостаточном восстановлении кислорода электронами. В результате, к кислороду могут присоединяться не 4, а сначала 1, или 2, или 3 электрона. При этом образуются активные формы кислорода: соответственно - сингле́тный кислоро́д, супероксид анион радикал, пероксид водорода, гидроксильный радикал. 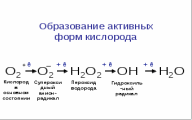
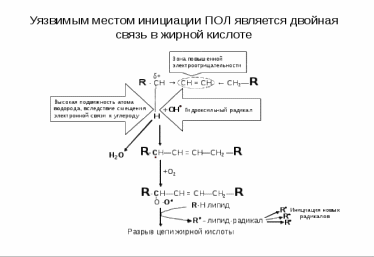 Стадии ПОЛ Стадии ПОЛ0) Образование активных форм кислорода. 1) Инициирование цепной реакции: в липидный слой мембран или липопротеинов внедряется свободный радикал, чаще всего HO• HO• способен приникать в толщу гидрофобного липидного слоя и вступать в химическое взаимодействие с полиненасыщенными жирными кислотами (LH), входящими в состав биологических мембран и липопротеинов плазмы крови. При этом образуются липидные радикалы: HO• + LH = H2O + L•. 2) Развитие цепи Липидный радикал (L• ·) вступает в реакцию с растворенным в среде молекулярным кислородом, при этом образуется новый свободный радикал – радикал липоперекиси (LOO·): . L• + O2 = LOO•. Этот радикал атакует одну из соседних молекул фосфолипида с образованием гидроперекиси липида LOOH и нового радикала L• ·: LOO• + LH = LOOH + L•. пероксид липида 3) Продолжение цепи: Образующиеся радикалы L•. инициируют новые цепи окисления липидов. L• + O2 = LOO•. 4) Обрыв цепи Цепь обрывается в результате взаимодействия свободных радикалов с антиоксидантами (InH), LOO• + InH = In•,: LOO• + Fe2+ + H˖ = LOOH. или друг с другом: LOO• + LOO• = молекулярные продукты 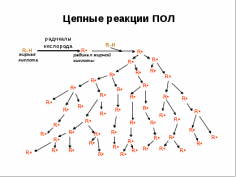 Глубина пероксидного окисления липидов зависит в первую очередь от состава липидов: с увеличением степени ненасыщенности жирных кислот, входящих в состав липидов, скорость окисления возрастает. Пероксидное окисление липидов приводит к повреждению клеточных мембран, структурными элементами которых являются липиды. Объектами ПОЛ могут служить химические связи липидов мембран, гиалуроновой кислоты, протеогликанов, коллагена. В присутствии ионов двухвалентного железа (Fе2+) АФК превращают оксигемоглобин в метгемоглобин. Окисляют функциональные группы белков, ферментов, гормонов и их рецепторов. Нарушают структуру азотистых оснований ДНК, кольца дезоксирибозы. Антиоксиданты — вещества, замедляющие или предотвращающие свободнорадикальные окислительные процессы. Антиоксидантами по отношению к ненасыщеным липидам являются витамины группы Е – токоферолы, которые ингибируют процесс пероксидного окисления. Антиокислительная функция токоферолов определяется наличием подвижного атома Н и способностью реагировать с активными радикалами с образованием малоактивных радикалов, стабилизированных р,p-сопряжение. Водорастворимые антиоксиданты: витамин С, тиолосодержащие соединения, растительные фенолы, Витамин С - восстанавливает окисленную форму витамина Е, взаимодействуя с водорастворимыми активными формами кислорода - Н2О2, ОН• инактивирует их. Глутатион - восстановитель, что позволяет ему взаимодействовать с различными соединениями-окислителями, такими как Н2О2, О2 • , ОН•-, с липидными пероксидами, дисульфидами, аскорбатом и другими, отдавая им атом водорода Жирорастворимые витамины: Е (Ток-ОН), Вит.А , Вит.К , молекулы антиоксиданта взаимодействуют с активными радикалами с образованием малоактивных радикалов; in vivo главным прерывающим цепь антиоксидантом является фермент супероксиддисмутаза, витамин Е, улавливающий свободные радикалы в липидной фазе. Витамин Е является наиболее широко распространенным антиоксидантом в природе. Он – один из самых важных жирорастворимых антиоксидантов в организме. Витамин E состоит из нескольких производных токоферолов. Главным его изомером в тканях человека является α-токоферол (α- Ток-ОН – образует малоактивный радикал (Ток-О• ) из сопряженных систем): α -Ток-ОН +R• → Ток-О• +RН |