сск. Структура светособирающих пигментных комплексов
 Скачать 231.92 Kb. Скачать 231.92 Kb.
|

|
Структура светособирающих пигментных комплексов Фотосинтез начинается с улавливания света пигментами –хлорофиллами входящими в состав хлоропластов клеток. Совокупность молекул светособирающего комплекса и реакционного центра составляет фотосистему. Предположение о существовании в хлоропластах двух фотосистем высказал Р. Эмерсон в 1957 г, изучая влияние света на квантовый выход (количество выделившегося кислорода или связанного углекислого газа на 1 квант поглощенной энергии ) фотосинтеза у водоросли хлореллы. Пигменты хлоропластов объединены в функциональные комплексы - пигментные системы, в которых реакционный центр (РЦ) хлорофилл а, осуществляющий фотосенсибилизацию, связан процессами переноса энергии с антенной, состоящей из светособирающих пигментов. Считают, что при низкой интенсивности света собственные антенны ФС I и ФС II поглощают мало квантов света и поэтому не могут поддерживать необходимой скорости фотосинтезирующих реакций, и нужен дополнительный приток энергии, который обеспечит ССК. Наоборот, при высоких интенсивностях света пигментным системам не нужно столько энергии и поэтому они содержат значительно меньше ССК. В состав ССК входят ксантофиллы, хлорофилл b, некоторое количество хлорофилла а. ССК передает поглощенную энергию на ПБК и затем на реакционный центр. Получив эту энергию, специальная форма хлорофилла в реакционном центре переходит в возбужденное состояние (Хл*). Хл* обладает очень высокой реакционной способностью и является сильным восстановителем. Современная схема фотосинтеза высших растений включает 2 фотохимические реакции, осуществляемые с участием 2 различных ФС. Предположение об их существовании было высказано Р. Эмерсоном в 1957 г. на основании обнаруженного им эффекта усиления действия длинноволнового (ДВ) красного света (700 нм) совместным освещением более коротковолновыми лучами (650 нм). Впоследствии с помощью обработки ультразвуком и поверхностно-активными веществами и использования дифференциального центрифугирования в градиенте плотности сахарозы удалось получить легкие фрагменты хлоропластов, обогащенные фракцией ФС1, и более тяжелые фрагменты, содержащие фракцию ФС2. Установлено, что ФС II поглощает более коротковолновые лучи по сравнению с ФС 1. Фотосинтез идет эффективно только при их совместном функционировании, что объясняет эффект усиления Эмерсона. ФС I: в качестве РЦ выступает димер хлорофилла а с максимумом поглощения света 700 нм (P700), роль антенного компонента играют также хлорофиллы а675-695. Первичным акцептором электронов в этой системе является мономерная форма хлорофилла а695, вторичными акцепторами - железосерные белки (-FeS). Комплекс ФС 1 под действием света восстанавливает железосодержащий белок - ферредоксин (Фд) и окисляет медьсодержащий белок- пластоцианин (Пц). ФC II: включает РЦ, содержащий хлорофилл а (P680) и антенные пигменты - хлорофиллы a670-683. Первичным акцептором электронов является феофитин (фф), передающий электроны на пластохинон. В состав ФС II входит также белковый комплекс S-системы, окисляющий воду, и переносчик электронов Z. Этот комплекс функционирует с участием Mn, Cl и Mg. ФС II восстанавливает пластохинон (PQ) и окисляет воду с выделением О2 и протонов. Связующим звеном между ФС II и ФС 1 служат фонд пластохинонов, белковый цитохромный комплекс (b6, f) и пластоцианин. На один РЦ приходится примерно 300 молекул пигментов, которые входят в состав антенных или светособирающих комплексов (ССК). Светособирающий белковый комплекс ламелл связан с ФСII и содержит хлорофиллы а, b и каротиноиды. В каждом ССК содержится от 120 до 240 молекул хлорофилла. Антенные комплексы входят в состав ФС1 и ФСII (фокусирующие антенные компоненты ФС). Антенный белковый комплекс ФС 1 содержит 110 молекул хлорофиллов а680-695 на один P700, из них 60 молекул - компоненты антенного комплекса, который можно рассматривать как ССК ФС1. Антенный комплекс ФС1 также содержит бета-каротин. Антенный белковый комплекс ФС II содержит 40 молекул хлорофиллов а670-683 на один P680 и бета-каротин. В окислительно-восстановительной системе А/А– происходит восстановление А до А–. От А– электрон переходит дальше и через другие компоненты электрон-транспортной цепи в конечном счете переносятся на НАДФ+. Хлорофилл, который отдал электрон, переходит в состояние свободного катиона – радикала Х+. Он возвращается в основное первичное состояние, получив электрон восстановленной формы Д– – второй окислительно-восстановительной системы Д/Д–. Окисленный Д восстанавливается до Д–, получая электрон от воды. 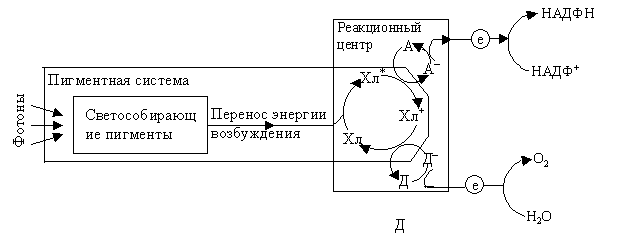 Рис. 2.11. Упрощенная схема работы пигментной системы Рис. 2.11. Упрощенная схема работы пигментной системыФС (II) и ФС (I) размещаются последовательно в цепи транспорта электронов от Н2О до НАДФ+. Для восстановления одной молекулы НАДФ+ в процессе фотосинтеза необходимо два электрона и два протона, при этом донором электронов является вода. Фотоиндуцированное окисление воды происходит в ФС II, восстановление НАДФ+ – ФС I. Таким образом, фотосистемы должны функционировать во взаимодействии. Хромопротеины антенных комплексов не обладают фотохимической активностью. Их роль состоит в поглощении и передаче энергии квантов на небольшое количество молекул РЦ P700 и P680, каждая из которых связана с цепью транспорта электронов и осуществляет фотохимическую реакцию. Организация электронно-транспортных цепей (ЭТЦ) при всех молекулах хлорофилла нерациональна, так как даже на прямом солнечном свету на молекулу пигмента кванты света попадают не чаще одного раза за 0,1 с. Передача энергии в светособирающих комплексах и ФС происходит путем индуктивного резонанса. Условиями для резонансного переноса энергии служат сближенное расположение и перекрытие частот осциллирующих электронных систем у двух взаимодействующих молекул. Причем миграция энергии осуществляется от коротковолновых пигментов к более длинноволновым. Квант света мигрирует в ФС, пока не попадет в стоковую молекулу (ловушку) - пигмент реакционного центра, который является самым длинноволновым и поэтому неспособным возбуждать соседние молекулы. Энергия возбужденного пигмента РЦ расходуется на фотохимическую работу. Квант света обычно передается на стоковую молекулу через несколько сотен передач, каждая передача осуществляется за 10-12 с, 100 передач-соответственно за 10-10 с. Экспериментально установлено, что 90 % энергии возбуждения хлорофилла а может быть передано на P700 менее чем за 10-9 с. Дело в том, что хотя передача энергии от одной молекулы пигмента к другой идет с большой эффективностью (от хлорофилла b к хлорофиллу а — 90%, от каротиноидов к хлорофиллу — 40%), однако все же это связано с некоторой ее потерей. Вместе с тем кванты света с меньшей длиной волны обладают большей энергией. Потеря энергии приводит к превращению квантов в более мелкие (с большей длиной волны). Именно поэтому основные формы хлорофилла, к которым стекается энергия, являются более длинноволновыми. Обратный перенос энергии невозможен. В том случае, если молекулы пигментов имеют одинаковый максимум поглощения возможен другой путь передачи энергии, получивший название экситонного. При этом пути электронное возбуждение распространяется в результате межмолекулярной вибрации на комплекс пигментов. Строение ССК зависит не только от систематического положения фотосинтезирующего организма, но и от условий произрастания. Теневыносливые растения имеют, как правило, больший размер ССК по сравнению с растениями, растущими в условиях высокой освещенности. Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза и заключаемая в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов. Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции. При поглощении фотона молекула хлорофилла переходит из основного (S0) в одно из возбужденных состояний – S1 или S2 с более высокой энергией. Возбужденное состояние S2 не стабильно и электрон в течение 10-12 сек. опускается на уровень S1 , теряя при этом часть энергии в виде тепла. Время пребывания электрона на S1 уровне составляет 10-9 сек. Он может изменить спин и перейти на Т1 уровень. Дезактивация S1 сопровождается следующими процессами: - превращение энергии возбуждения в тепло (релаксация); - излучение кванта с переходом электрона на S0 уровень (флуоресценция или фосфоресценция); - перенос световой энергии на другую молекулу пигмента; - использование энергии возбуждения в фотохимической реакции. 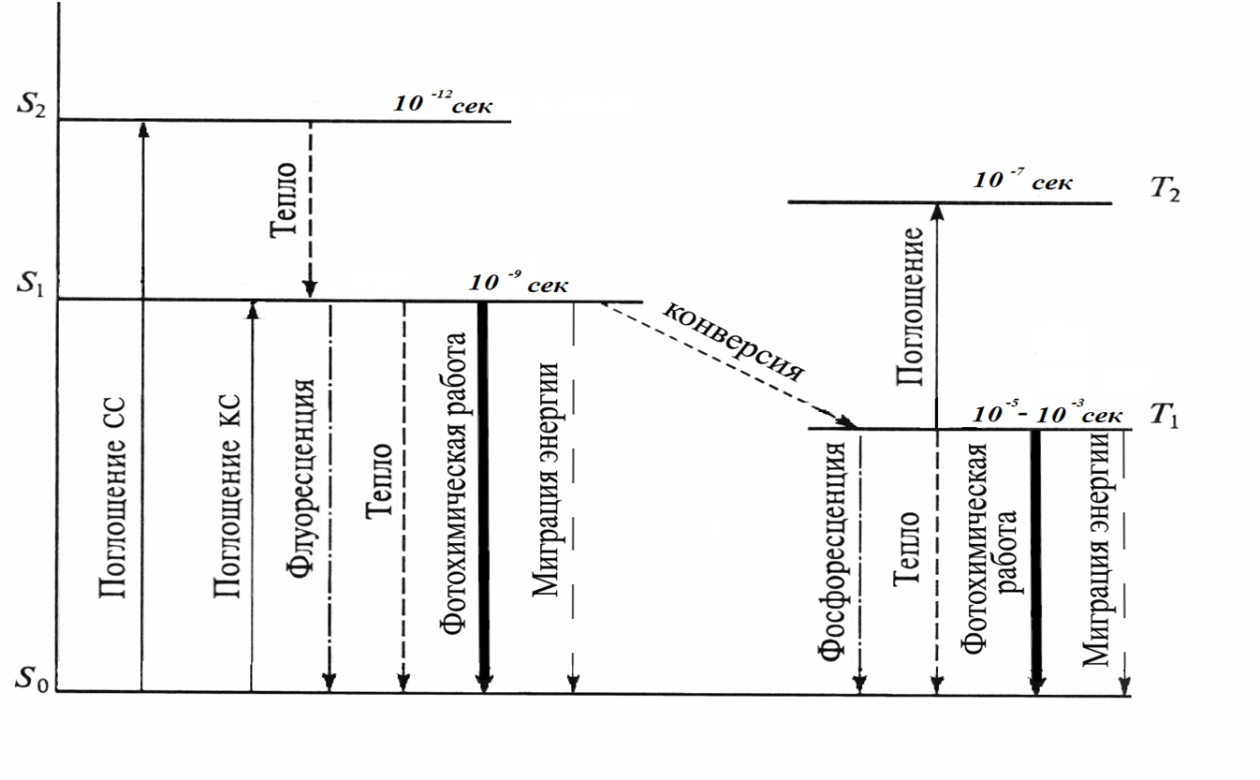 |