Свойства возбудимых тканей. Биоэлектричество Понятие раздражимости и раздражителя. Классификация раздражителей, понятие адекватного раздражителя
 Скачать 6.69 Mb. Скачать 6.69 Mb.
|

|
Физиология мышц. 1. Функции мышечной ткани в организме. Свойства и функции поперечнополосатых мышц. Особенности гладких мышц. Одним из наиболее распространенных проявлений возбуждения в возбудимых тканях является изменение длины — сокращение. В основе способности к изменению длины лежит свойство сократимости, наиболее выраженное в мышечной ткани. Различают поперечнополосатую и гладкую мышечную ткань. Гладкая мышечная ткань представлена во внутренних полых органах: мышечном слое стенки сосудов, желудочно-кишечного тракта, мочевого пузыря, матки и некоторых других. Поперечнополосатая мышечная ткань составляет основу скелетных мышц. Особую разновидность поперечнополосатой мускулатуры представляет сердечная мышца. Физиологические особенности сердечной мышцы будут рассмотрены при изучении системы кровообращения. ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ ОПОРНО- ДВИГАТЕЛЬНОГО АППАРАТА Любая скелетная мышца состоит из трех типов мышечных волокон: ▪ быстрых, фазических (в связи с относительно низким содержанием пигмента миоглобина их называют белыми); ▪ медленных, тонических (в связи с относительно высоким содержанием окрашивающего миоглобина их называют красными); ▪ волокон-рецепторов. 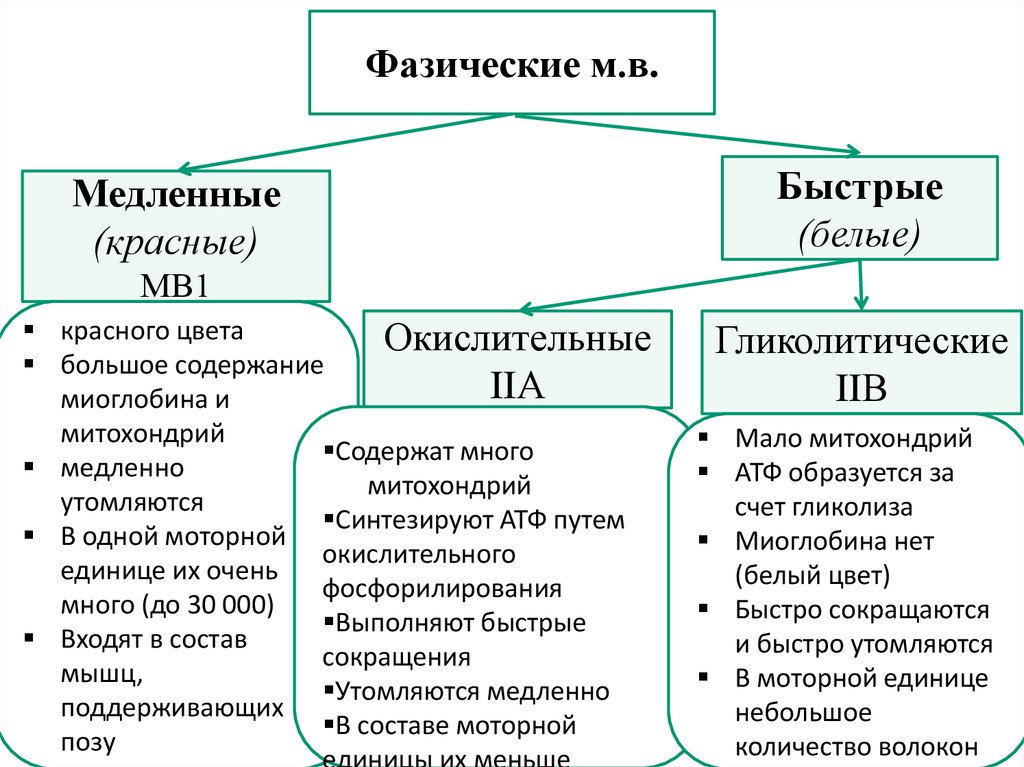 Основная функция первых двух групп волокон состоит в изменении длины мышц: формирование мышечного тонуса и различных вариантов быстрых (фазических) сокращений. Первые две группы мышечных волокон объединяют в группу экстрафузальных мышечных волокон. У волокон-рецепторов есть название-синоним — интрафузальные мышечные волокна. Рассмотрим особенности строения и свойств экстрафузальных волокон. Выделяют физические и физиологические свойства мышечной ткани. К физическим свойствам мышечной ткани относят вязкость, эластичность, пластичность. К физиологическим — раздражимость, возбудимость, лабильность, проводимость.   ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ ● Растяжимость — способность менять длину под влиянием приложенной силы. ● Эластичность — способность мышцы восстанавливать первоначальную форму после прекращения действия сил, вызывающих ее деформацию. ● Сила мышц — максимальный груз, который мышца способна приподнять. ● Работа мышц — произведение поднимаемого груза на высоту подъема, измеряется в килограммах на метр (кг/м): А = Р × Δh, где А — работа, выполненная мышцей; Р — масса груза; Δh — высота, на которую поднят груз. Максимальный объем выполненной мышцей работы отмечается при средних величинах нагрузки   СТРОЕНИЕ СОМАТИЧЕСКОЙ ПОПЕРЕЧНОПОЛОСАТОЙ МЫШЕЧНОЙ ТКАНИ С 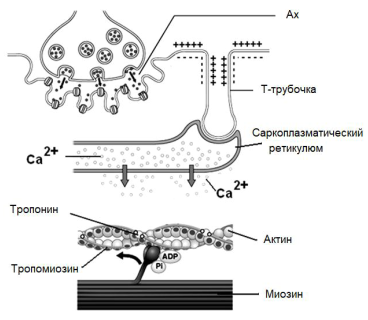 келетная поперечнополосатая мышечная ткань состоит из мышечных волокон. Каждое мышечное волокно содержит до 1000, а в ряде случаев и более сократительных волокон — миофибрилл. Каждая миофибрилла состоит из множества толстых и тонких нитей — миофиламентов (соотношение тонких и толстых нитей 2:1). Каждое мышечное волокно поперечнополосатой мускулатуры содержит большое количество ядер и представляет собой симпласт. Толстые нити состоят из белка миозина, а тонкие — из актина. Кроме того, в состав тонких нитей входят еще белки тропонин и тропомиозин, формирующие с актомиозином единый регуляторный комплекс. Миозиновые нити имеют поперечно ориентированные мостики, которые участвуют в механизмах взаимодействия актиновых и миозиновых нитей (рис. 5.2). келетная поперечнополосатая мышечная ткань состоит из мышечных волокон. Каждое мышечное волокно содержит до 1000, а в ряде случаев и более сократительных волокон — миофибрилл. Каждая миофибрилла состоит из множества толстых и тонких нитей — миофиламентов (соотношение тонких и толстых нитей 2:1). Каждое мышечное волокно поперечнополосатой мускулатуры содержит большое количество ядер и представляет собой симпласт. Толстые нити состоят из белка миозина, а тонкие — из актина. Кроме того, в состав тонких нитей входят еще белки тропонин и тропомиозин, формирующие с актомиозином единый регуляторный комплекс. Миозиновые нити имеют поперечно ориентированные мостики, которые участвуют в механизмах взаимодействия актиновых и миозиновых нитей (рис. 5.2). Наряду с миофибриллами в саркоплазме мышечных волокон имеется система канальцев — саркоплазматический ретикулум, которые имеют расширения в области мембраны мышечного волокна (цистерны саркоплазматического ретикулума). В цистернах саркоплазматического ретикулума депонируются ионы кальция — Са++. Мембрана мышечного волокна в области прилежания цистерн имеет характерную форму — форму буквы Т (см. рис. 5.2). Поскольку участок мембраны саркоплазмы и цистерны саркоплазматического ретикулума связаны единой функцией, этот комплекс получил название Т-системы. ОСОБЕННОСТИ ГЛАДКОМЫШЕЧНОЙ ТКАНИ Гладкомышечные клетки распределены в стенках внутренних органов — сосудах, органах желудочно-кишечного тракта, мочевом пузыре, мочевыводящих путях, матке. Гладкомышечные клетки имеют веретенообразную форму длиной и содержат одно ядро. Сократительные элементы так же, как и в скелетной мускулатуре, представлены миофиламентами, содержащими актин, миозин и регуляторные белки. Однако, в отличие от скелетной мускулатуры, в гладкомышечной ткани миофиламенты расположены беспорядочно, поэтому нет поперечной исчерченности. В гладкомышечных клетках слаборазвит саркоплазматический ретикулум, поэтому ионы Са++ депонируются в цитоплазме клеток. Особенность миозина гладкомышечной ткани — низкая АТФазная активность, что приводит к медленному скольжению нитей актина и миозина вдоль друг друга. В результате гладкомышечная ткань характеризуется медленной, тонической формой мышечного сокращения. Кроме того, это приводит к низкой энергозатратности сокращения, поэтому в гладкомышечной ткани утомление развивается медленнее, чем в поперечнополосатой ткани скелетных мышц. Между отдельными клетками гладкомышечной ткани имеются специальные образования — нексусы (разновидности электрических синапсов), позволяющие возбуждению в виде биоэлектрического процесса достаточно быстро распространяться по ткани. Следует добавить, что гладкомышечные клетки способны переходить из состояния относительного покоя в состояние возбуждения без внешних воздействий, то есть обладают свойством автоматии.      2. Типы, формы и режимы сокращения мышц. Одиночное сокращение, тетанус, тонус, контрактура. Их анализ. Выделяют три режима (типа) мышечного сокращения: ▪ изотонический; ▪ изометрический; ▪ смешанный (ауксометрический). Изотонический режим мышечного сокращения характеризуется преимущественным изменением длины мышечного волокна без существенного изменения напряжения. Этот режим мышечного сокращения наблюдается, например, при поднятии легких и средних по массе грузов. Изометрический режим мышечного сокращения характеризуется преимущественным изменением мышечного напряжения, без существенного изменения длины. Примером могут служить изменения состояния мышц при попытке человека сдвинуть с места предмет большой массы (например, при попытке сдвинуть с места стену в комнате). Смешанный (ауксометрический) тип мышечного сокращения, наиболее реальный, наиболее часто встречающийся вариант. Содержит в себе компоненты первого и второго вариантов в разных соотношениях в зависимости от реальных условий окружающей среды. Выделяют три вида мышечного сокращения: ▪ одиночное; ▪ тетаническое (тетанус); ▪ тоническое. Кроме того, тетаническое мышечное сокращение делят на зубчатый и гладкий тетанус.  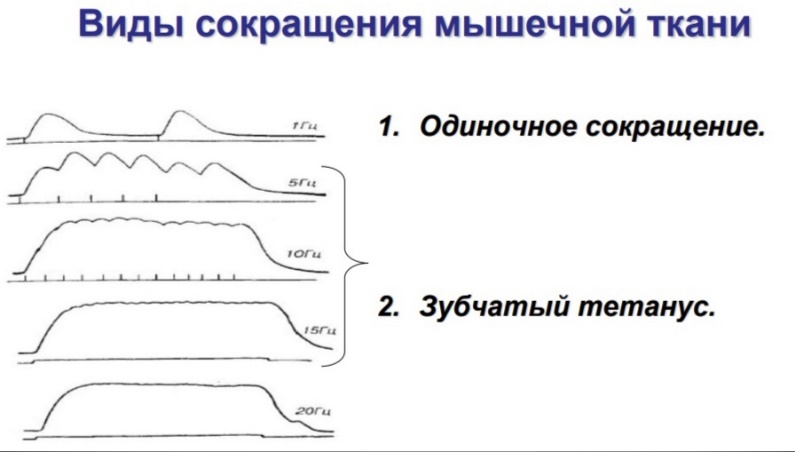 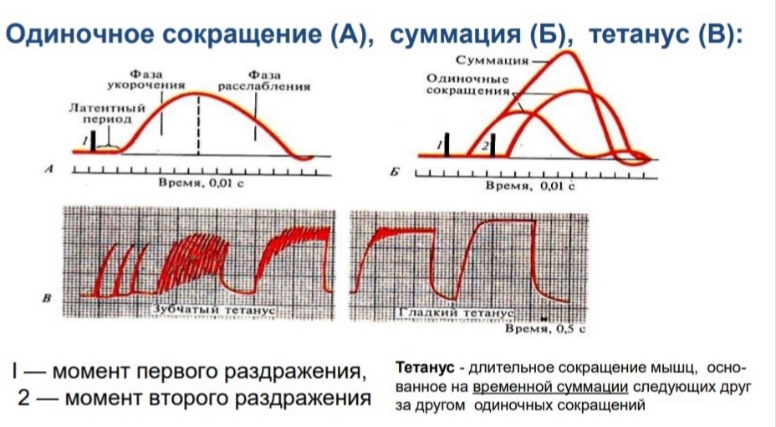 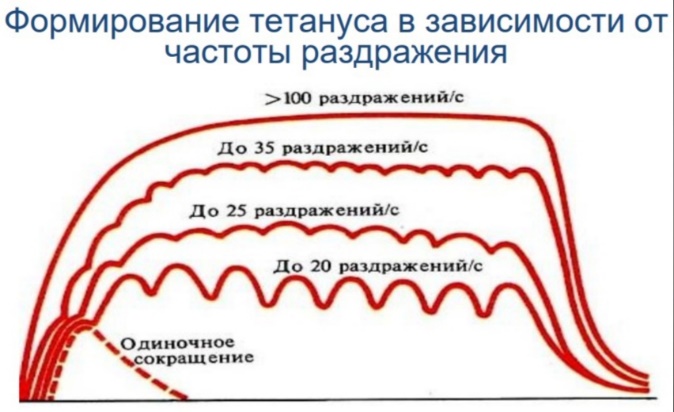  О 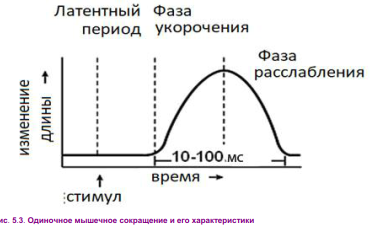 диночное мышечное сокращение возникает в условиях действия на мышцу пороговых или надпороговых электрических стимулов, межимпульсный интервал которых равен или больше длительности одиночного мышечного сокращения. В одиночном мышечном сокращении выделяют три временных отрезка: латентный период, фазу укорочения и фазу расслабления (рис. 5.3). диночное мышечное сокращение возникает в условиях действия на мышцу пороговых или надпороговых электрических стимулов, межимпульсный интервал которых равен или больше длительности одиночного мышечного сокращения. В одиночном мышечном сокращении выделяют три временных отрезка: латентный период, фазу укорочения и фазу расслабления (рис. 5.3). Т 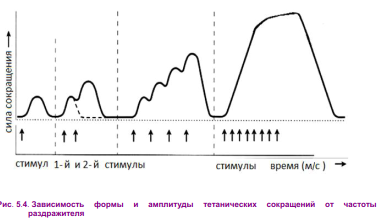 етаническое мышечное сокращение (тетанус) возникает в условиях действия на скелетную мышцу порогового или надпорогового электрического раздражителя, межимпульсный интервал которого меньше длительности одиночного мышечного сокращения. В зависимости от длительности межстимульных интервалов электрического раздражителя при его воздействии может возникнуть либо зубчатый, либо гладкий тетанус. етаническое мышечное сокращение (тетанус) возникает в условиях действия на скелетную мышцу порогового или надпорогового электрического раздражителя, межимпульсный интервал которого меньше длительности одиночного мышечного сокращения. В зависимости от длительности межстимульных интервалов электрического раздражителя при его воздействии может возникнуть либо зубчатый, либо гладкий тетанус. Если межимпульсный интервал электрического раздражителя меньше длительности одиночного мышечного сокращения, но больше или равен сумме латентного периода и фазы укорочения, возникает зубчатый тетанус. Это условие выполняется при повышении частоты импульсного электрического раздражителя в определенном диапазоне. Если же длительность межимпульсного интервала электрического раздражителя меньше суммы латентного периода и фазы укорочения, возникает гладкий тетанус. При этом амплитуда гладкого тетануса больше амплитуды и одиночного мышечного сокращения, и зубчатого тетанического сокращения. При дальнейшем уменьшении межимпульсного интервала электрического раздражителя, а следовательно, при увеличении частоты амплитуда тетанических сокращений возрастает (рис. 5.4). О 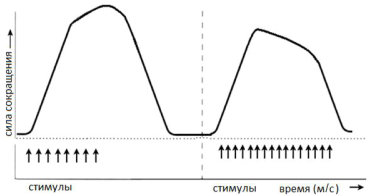 днако эта закономерность не носит абсолютного характера: при определенном значении частоты вместо ожидаемого повышения амплитуды гладкого тетануса отмечается феномен ее снижения (рис. 5.5). Этот феномен был впервые обнаружен российским ученым Н.Е. Введенским и назван пессимумом. В основе пессимальных явлений, по мнению Н.Е. Введенского, лежит механизм торможения, возникающий при условии превышения частоты предъявляемого раздражителя, меры лабильности мышечной ткани. днако эта закономерность не носит абсолютного характера: при определенном значении частоты вместо ожидаемого повышения амплитуды гладкого тетануса отмечается феномен ее снижения (рис. 5.5). Этот феномен был впервые обнаружен российским ученым Н.Е. Введенским и назван пессимумом. В основе пессимальных явлений, по мнению Н.Е. Введенского, лежит механизм торможения, возникающий при условии превышения частоты предъявляемого раздражителя, меры лабильности мышечной ткани. Рис. 5.5. Зависимость амплитуды гладкого тетануса от частоты раздражителя МЕХАНИЗМЫ ТЕТАНИЧЕСКОГО СОКРАЩЕНИЯ Одна из наиболее распространенных теорий тетанического сокращения предложена в XIX в. известным немецким физиологом Г. Гельмгольцем. Эта теория получила название «теория суперпозиции» («теории суммации»). Согласно представлениям Г. Гельмгольца, тетанус возникает в результате суммации механических проявлений изменения длины скелетных мышц. Если суммация последующего изменения длины мышцы с предыдущим происходит в условиях, когда мышечное волокно находится в фазе расслабления, возникает зубчатый тетанус. Если же суммация происходит в условиях попадания каждого последующего стимула на мышечное волокно, находящееся в фазе укорочения, возникает гладкий тетанус (рис. 5.6). 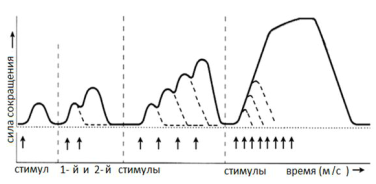 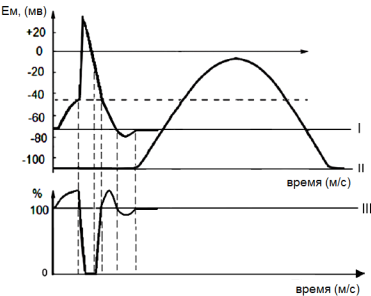 Рис. 5.6. Суммация мышечных сокращений при возникновении тетануса Рис. 5.7. Соотношение между изменением длины мышечного волокна в условиях одиночного мышечного сокращения, изменением мембранного потенциала и возбудимости Физиологическая основа возможности суммации мышечных сокращений при действии относительно высокочастотных электрических раздражителей становится ясной при рассмотрении соотношений между изменением длины мышечного волокна в условиях одиночного мышечного сокращения, изменением мембранного потенциала и возбудимости мышцы (рис. 5.7). Как следует из представленных соотношений, колебание мембранного потенциала при возбуждении мышечного волокна завершается в течение латентного периода одиночного мышечного сокращения. К этому же моменту мышечное волокно полностью восстанавливает свою возбудимость. В связи с этим оно способно полноценно возбуждаться, проявлением чего становится полноценное дополнительное изменение длины мышечного волокна, которое может суммироваться с предыдущим. 3. Двигательные единицы и их характеристика. Поперечнополосатая скелетная мышечная ткань иннервируется соматической нервной системой. Нейроны соматической нервной системы, иннервирующие скелетную мускулатуру, называются мотонейронами. Мотонейроны, иннервирующие белые мышечные волокна, называются альфа-1 мотонейронами (α1-мотнейроны), иннервирующие красные мышечные волокна, — альфа-2 мотонейронами (α2-мотнейроны). Один мотонейрон, как правило, иннервирует достаточно много мышечных волокон (от 12 до 2500). Вместе с иннервирующим их мотонейроном мышечные волокна составляют двигательную или нейромоторную единицу. Двигательные единицы, участвующие в обеспечении тонких движений, содержат от 12 до 15 мышечных волокон. К таким нейромоторным единицам относятся, например, двигательные единицы мышц кистей рук. Двигательные единицы, участвующие в обеспечении грубых движений, содержат от 1500 до 2500 мышечных волокон. К таким двигательным единицам относят, например, нейромоторные единицы мышц нижних конечностей, глубоких мышц спины. В соответствии с составом двигательных единиц их делят на три группы: ▪ I — медленные, неутомляемые двигательные единицы; ▪ IIА — быстрые, устойчивые к утомлению двигательные единицы; ▪ IIБ — быстрые, легко утомляемые двигательные единицы. Медленные, неутомляемые двигательные единицы состоят из мелких α2- мотонейронов и красных тонических мышечных волокон. Характеризуются высокой возбудимостью, то есть переходят в состояние возбуждения при относительно низкой частоте разрядов ПД нервных клеток. Красные мышечные волокна, входящие в состав данных двигательных единиц, характеризуются хорошо развитой капиллярной сетью, что позволяет им поддерживать длительное возбуждение. Быстрые_,_устойчивые_к_утомлению_двигательные_единицы'>Быстрые, устойчивые к утомлению двигательные единицы состоят из α1- мотонейронов и белых мышечных волокон. Как и единицы в предыдущей группе, характеризуются хорошо развитой капиллярной сетью. Мышечные волокна для воспроизводства энергии используют как аэробные, так и анаэробные процессы. По уровню возбудимости относятся к группе средневозбудимых двигательных единиц. Быстрые, легко утомляемые двигательные единицы состоят из α1- мотонейронов и белых мышечных волокон. Из представленных трех групп двигательных единиц характеризуются самой низкой возбудимостью и вместе с тем максимальной амплитудой мышечного сокращения. Биохимическая особенность данной группы — способность мышечных волокон извлекать энергию для своих нужд за счет анаэробных процессов, то есть без участия кислорода. Из представленной выше классификации следует, что нейромоторные или двигательные единицы обладают неодинаковой возбудимостью, поэтому при увеличении силы раздражителя количество вовлеченных в процесс возбуждения двигательных единиц увеличивается, что закономерно приводит к увеличению амплитуды мышечного сокращения. Данная закономерность называется законом «силы». Он формулируется следующим образом: «амплитуда сокращения скелетных мышц зависит от силы раздражителя: в определенном диапазоне с увеличением силы раздражителя возрастает амплитуда мышечного сокращения». 4. Работа и сила мышц. Величина сокращения (степень укорочения) мышцы при данной силе раздражения (т. е. при данном числе активированных волокон) зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект; при сильном растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения снижается. Для измерения силы мышцы определяют либо тот максимальный груз, который она в состоянии поднять, либо максимальное напряжение, которое она может развить в условиях изометрического сокращения. Эта сила может быть очень велика. Установлено, что собака мышцами челюсти может поднять груз, превышающий массу ее тела в 8,3 раза. Одиночное мышечное волокно способно развить напряжение 100—200 мг. Учитывая, что общее количество мышечных волокон в теле человека приблизительно 15—30 млн., они могли бы развить напряжение в 20—30 т, если бы все одновременно тянули в одну сторону. Сила мышцы при прочих равных условиях зависит от ее поперечного сечения. Чем больше физиологическое поперечное сечение мышцы, т. е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон сумма поперечных сечений может значительно превышать геометрическое поперечное сечение самой мышцы (рис. 36). По этой причине сила мышцы с косо расположенными волокнами значительно больше, чем сила мышцы той же толщины, но при продольном расположении волокон. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который в состоянии поднять мышца, делят на площадь ее физиологического поперечного сечения. Таким образом вычисляют абсолютную мышечную силу. Абсолютная сила, выраженная в килограммах на 1 см2, икроножной мышцы человека равна 5,9, сгибателя плеча — 8,1, жевательной мышцы — 10, двуглавой мышцы плеча — 11,4, трехглавой мышцы плеча — 16,8, гладких мышц— 1. Примером мышц с продольным расположением волокон может служить m.sartorius, с косым — m.m.intercostales. Большинство мышц млекопитающих и человека перистого строения. Перистая мышца имеет большое физиологическое сечение, а потому обладает большой силой.    |