Свойства возбудимых тканей. Биоэлектричество Понятие раздражимости и раздражителя. Классификация раздражителей, понятие адекватного раздражителя
 Скачать 6.69 Mb. Скачать 6.69 Mb.
|

|
5. Механизм мышечного сокращения и расслабления: роль ПД, ионов Са, АТФ. МЕХАНИЗМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ Структура миофибрилл и ее изменения при сокращении. Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими' свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением.В обыкновенном свете они выглядят темными, а в поляризованном — прозрачными в продольном направлении и непрозрачными в поперечном. Другие участки в обыкновенном свете выглядят светлыми — они изотропны, т. е. не обладают двойным лучепреломлением (рис. 34, а). Анизотропные участки обозначают буквой Л, изотропные — буквой I. В середине диска Л различается светлая полоска Н, посередине диска I — темная полоска Z, представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрнлл внутри одного волокна во время сокращения не смещаются по отношению друг к другу. Современные представления о структуре миофибриллярпого аппарата основываются на исследованиях структуры мышечного волокна при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами. Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимери-зованные молекулы белков миозина и актина. Миозиновые протофибриллы, или, как их принято обозначать, нити, вдвое толще актиповых. Их диаметр примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски Л — из нитей миозина, а, возможно, еще и другого белка. Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. Мембрана Z, проходя через середину диска I, скрепляет между собой эти нити: Важным компонентом ультрамикроскопической структуры миофибрилл являются также многочисленные поперечные мостики, соединяющие между собой миозиновые и актиновые нити. При сокращении мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски I укорачиваются, а диски Л сохраняют свой размер. Почти исчезает светлая полоска Н, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствий-ионов Са24 и АТФ. Наблюдается своего рода химическое «зубчатое колесо», как бы протягивающее одну группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей. Теория сокращения: при сокращении происходит уменьшение размера саркомера вследствие активного перемещения тонких актиновых нитей относительно толстых миозиновых. Механизм мышечного сокращения. В процессе сокращения мышечного волокна в нем происходят следующие преобразования: А. Электрохимическое преобразование: 1. Генерация ПД. 2. Распространение ПД по Т-системе. 3. Электрическая стимуляция зоны контакта Т-системы и саркоплазматического ретикулума, активация ферментов, образование инозитолтрифосфата, повышение внутриклеточной концентрации ионов Са2+. Б. Хемомеханическое преобразование: 4. Взаимодействие ионов Са2+ с тропонином, освобождение активных центров на актиновых филаментах. 5. Взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги. 6. Скольжение нитей актина и миозина относительно друг друга, уменьшение размера саркомера, развитие напряжения или укорочение мышечного волокна. Передача возбуждения с двигательного мотонейрона на мышечное волокно происходит с помощью медиатора ацетилхолина (АХ). Взаимодействие АХ с холинорецептором концевой пластинки = активации АХ-чувствительных каналов и появлению потенциала концевой пластинки. При этом область концевой пластинки становится источником раздражающего тока для мембраны мышечного волокна и на участках клеточной мембраны, прилегающих к концевой пластинке, возникает ПД, который распространяется в обе стороны. Таким образом, генерация ПД- 1ый этап МС. Второй этап: распространение ПД внутрь мышечного волокна по поперечной системе трубочек. Трубочки связывают поверхностную мембрану и сократительный аппарат мышечного волокна: Т-система тесно контактирует с терминальными цистернами саркоплазматической сети двух соседних саркомеров. Третий этап: Электрическая стимуляция места контакта приводит к активации ферментов и к образованию инозитолтрифосфата. Инозитолтрифосфат активирует кальциевые каналы мембран терминальных цистерн, что приводит к выходу ионов Са2+ из цистерн и повышению внутриклеточной концентрации Са2+. Таким образом, на первых этапах происходит преобразование электрического сигнала ПД в химический — повышение внутриклеточной концентрации Са2+, т. е. электрохимическое преобразование. При повышении внутриклеточной концентрации ионов Са2+ тропомиозин смещается в желобок между нитями актина, при этом на актиновых нитях открываются участки, с которыми могут взаимодействовать поперечные мостики миозина. Это смещение тропомиозина обусловлено изменением конформации молекулы белка тропонина при связывании Са2+-4ый этап 5ый этап: присоединение головки поперечного мостика к актиновому филаменту к первому из нескольких последовательно расположенных стабильных центров. При этом миозиновая головка поворачивается вокруг своей оси, поскольку имеет несколько активных центров, которые последовательно взаимодействуют с соответствующими центрами на актиновом филаменте. Вращение головки приводит к увеличению упругой эластической тяги шейки поперечного мостика и увеличению напряжения. В каждый конкретный момент в процессе развития сокращения одна часть головок поперечных мостиков находится в соединении с актиновым филаментом, другая свободна, т. е. существует последовательность их взаимодействия с актиновым филаментом. Это обеспечивает плавность процесса сокращения. На четвертом и пятом этапах происходит хемомеханическое преобразование. Последовательная реакция соединения и разъединения головок поперечных мостиков с актиновым филаментом приводит к скольжению тонких и толстых нитей относительно друг друга и уменьшению размеров саркомера и общей длины мышцы, что является шестым этапом. Первоначально полагали, что ионы Са2+ служат кофактором АТФазной активности миозина. Дальнейшие исследования опровергли это предположение. У покоящейся мышцы актин и миозин практически не обладают АТФазной активностью. Присоединение головки миозина к актину приводит к тому, что головка приобретает АТФазную активность. Гидролиз АТФ в АТФазном центре головки миозина сопровождается изменением конформации последней и переводом ее в новое, высокоэнергетическое состояние. Повторное присоединение миозиновой головки к новому центру на актиновом филаменте вновь приводит к вращению головки, которое обеспечивается запасенной в ней энергией. В каждом цикле соединения и разъединения головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Быстрота вращения определяется скоростью расщепления АТФ. Таким образом, в процессе хемомеханического преобразования АТФ разъединяет головки миозина и актинового филамента и обеспечивает энергетику для дальнейшего взаимодействия головки миозина с другим участком актинового филамента.. Для расслабления нужно понизить концентрацию ионов Са2+: саркоплазматическая сеть имеет кальциевый насос, который возвращает кальций в цистерны. Активация кальциевого насоса осуществляется неорганическим фосфатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса осуществляетс за счет энергии, образующейся при гидролизе АТФ. Таким образом, АТФ является вторым важнейшим фактором, абсолютно необходимым для процесса расслабления. . Нервно-мышечная передача. Строение синапса, механизм проведения возбуждения.  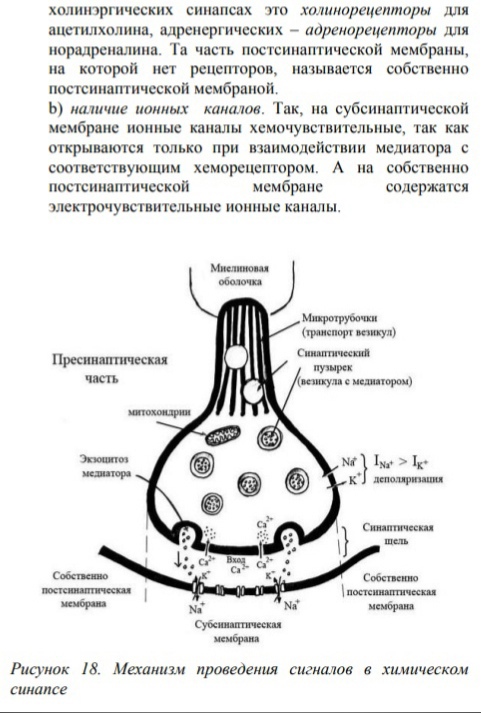       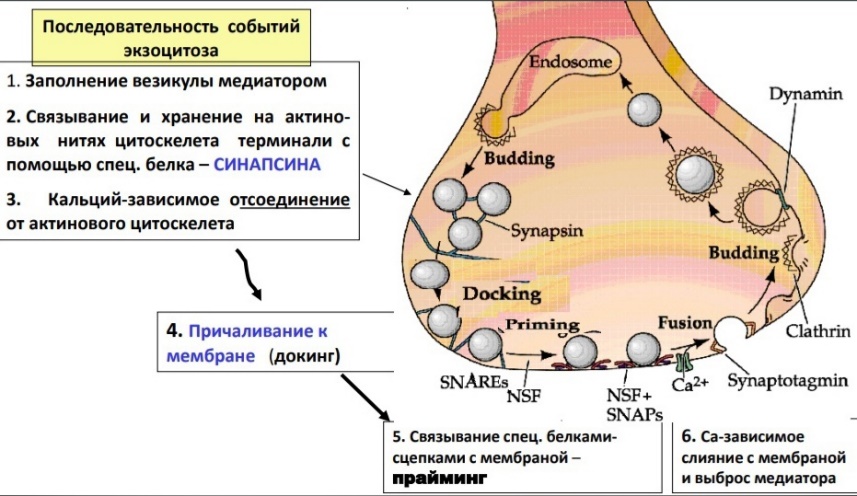 2. Пессимальное торможение: оптимум и пессимум, парабиоз, его фазы. 3. Механизм пессимального торможения, роль ритма раздражения и меры лабильности ткани.   4. Утомление как состояние ткани, органа, системы органов и целого организма. Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха. Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления. Утомление — процесс фазный, как и многие другие процессы в организме человека. В первой фазе возникает некоторое напряжение в деятельности физиологических систем. Устойчивое состояние может еще не нарушиться, но поддерживать его становится все труднее. Только современные математические приемы обработки результатов физиологических исследований с помощью компьютеров позволили «увидеть» эту фазу утомления. Во второй фазе уже отчетливо видны нарушения устойчивого состояния. При мышечной деятельности это проявляется в несогласованном снижении одних показателей и повышении других. Например, потребление кислорода может начать снижаться, а объемная скорость дыхания при этом возрастать. Это явный признак снижения эффективности и разбалансировки в деятельности вегетативных систем, характерный для утомления. При этом работа по-прежнему выполняется в том же объеме, с прежней интенсивностью: компенсаторные механизмы все еще справляются с удержанием необходимых функциональных свойств мышц. При умственной работе эта фаза обычно проявляется в увеличении числа ошибок, т.е. опять же в снижении эффективности, при сохранении скорости работы. Третья фаза — срыв устойчивого состояния. Разбалансировка в работе вегетативных систем быстро нарастает, их эффективность резко падает, и вслед за этим наступает отказ от работы («Не могу!»). Умственная работа, не требующая столь больших энергетических ресурсов, может при этом и продолжаться, однако ее неэффективность делает ее продолжение совершенно бессмысленным. Таким образом, чем дольше не наступает утомление при определенном уровне нагрузки либо чем выше уровень нагрузки, при котором наступает утомление, тем выше работоспособность человека. 5. Признаки утомления мышц, нерва, нервно-мышечного синапса. Причины утомления. Признаки утомления: Мышцы: 1. Увеличение латентного периода (отрезок времени от момента начала раздражения мышцы до момента начала сокращения). 2. Понижение возбудимости. Наряду с изменением амплитуды сокращения при утомлении нарастает латентный период сокращения и увеличиваются пороги раздражения и хронаксия, то есть понижается возбудимость. Эти изменение возникают не сразу после работы, а спустя некоторое время, в течение которого наблюдается увеличение амплитуды одиночных сокращений мышцы. Этот период называется периодом врабатывания. При дальнейшем длительном раздражении развивается утомление мышечных волокон. 3. Повышение вязкости мышцы. Нерва и нервно-мышечного аппарата: Нервно-мышечный препарат содержит в себе три элемента: мышечное волокно, нервно-мышечный синапс и нервное волокно. Опыт показывает, что при утомлении нервно-мышечного препарата изменение функциональных свойств наступает, в первую очередь, в нервно-мышечных синапсах, во вторую очередь, — непосредственно в мышечных волокнах. Что касается нервных проводников, то они, как впервые показал Н. Е. Введенский, практически «неутомимы». Изменение функциональных свойств нервно-мышечных синапсов выражается в нарушении процесса передачи возбуждения с нервных волокон на мышечные. Причины утомления: Мышцы: Понижение работоспособности изолированной из организма мышцы при ее длительном раздражении обусловлено двумя основными причинами: 1. первой из них является то, что во время сокращений в мышце накапливаются продукты обмена веществ (в частности, молочная, фосфорная кислоты и т. д.), оказывающие угнетающее влияние на работоспособность мышцы. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия. Если изолированную мышцу, помещенную в раствор Рингера, довести длительным раздражением до полного утомления, то достаточно только сменить омывающую ее жидкость, чтобы восстановить сокращения мышцы. 2. Другой причиной развития утомления изолированной мышцы является постепенное истощение в ней энергетический запасов. При длительной работе изолированной мышцы происходит резкое уменьшение запасов гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для осуществления сокращения. Необходимо отметить, что утомление изолированной скелетной мышцы при ее прямом раздражении является лабораторным феноменом. В естественных условиях утомление двигательного аппарата при длительной работе развивается более сложно и зависит от большего числа факторов. 1. В организме мышца непрерывно снабжается кровью, и, следовательно, получает с ней определенное количество питательных веществ (глюкоза, аминокислоты) и освобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. 2. В целом организме утомление зависит не только от процессов в мышце, но и от процессов, развивающихся в нервной системе, участвующих в управление двигательной деятельностью. Так, например, утомление сопровождается дискоординацией движений, возбуждением многих мышц, которые не участвуют в совершении работы. Нерва и нервно-мышечного аппарата: Утомление нервно-мышечного препарата обусловлено следующими причинами: 1. При длительном раздражении нерва нарушение нервно-мышечной передачи развивается задолго до того, как мышца, а тем б проведению возбуждения. Объясняется это тем, что в нервных окончаниях при длительном раздражении уменьшается запас "заготовленного" медиатора. Поэтому порции ацетилхолина, выделяющиеся в синапсах в ответ на каждый импульс, уменьшаются и постсинаптические потенциалы снижаются до подпороговых величин. 2. Наряду с этим при длительном раздражении нерва происходит постепенное понижение чувствительности постсинаптической мембраны мышечного волокна к ацетилхолину. В результате уменьшается величина потенциалов концевой пластинки. Когда их амплитуда падает ниже некоторого критического уровня, возникновение потенциалов действия в мышечном волокне прекращается. По этим причинам синапсы быстрее утомляются, чем нервные волокна и мышцы. Следует отметить, что нервные волокна обладают относительной неутомляемостью. Впервые Н.Е. Введенский показал, что нерв в атмосфере воздуха сохраняет способность к проведению возбуждений даже при многочасовом непрерывном раздражении (около 8 часов). Относительная неутомляемость нерва отчасти зависит от того, что нерв тратит при своем возбуждении сравнительно мало энергии. Благодаря этому процессы ресинтеза в нерве в состоянии покрывать его относительно малые расходы при возбуждении даже в том случае, если это возбуждение длится много часов. ТЕСТ (ДОПУСК) 1. Отсоединение головки миозина от актиновой нити - СВОБОДНОЙ АТФ 2. Структурное образование, обеспечивающее передачу возбуждения с одной клетки на другую, носит название – синапс 3. Определите верны или неверны утверждения и связь между ними Пессимум сокращения мышцы возникает при очень большой частоте раздражения, потому что при такой частоте каждый последующий импульс приходит в рефрактерные фазы от предыдущего – ВВВ 4. Медиатором в нервно-мышечном синапсе скелетных мышц человека является - |